By
Dr. suhair majeed

Cartilage is a special form of connective tissue that
also develops from the mesenchyme.
Similar to the connective tissue, cartilage consists
of
cells and extracellular matrix
composed of
connective tissue fibers and ground substance.
In contrast to connective tissue, cartilage is
nonvascular (avascular) and receives its nutrition
via diffusion through the extracellular matrix.

Cartilage exhibits tensile strength, provides
firm structural support for soft tissues, allows
flexibility without distortion, and is resilient to
compression.
Cartilage consists mainly of cells called
chondrocytes and chondroblasts that synthesize
the extensive extracellular matrix.
There are three main types of cartilage in the
body:
hyaline, elastic, and fibrocartilage
.
Their classification is based on the amount and
types of connective tissue fibers that are present
in the extracellular matrix.

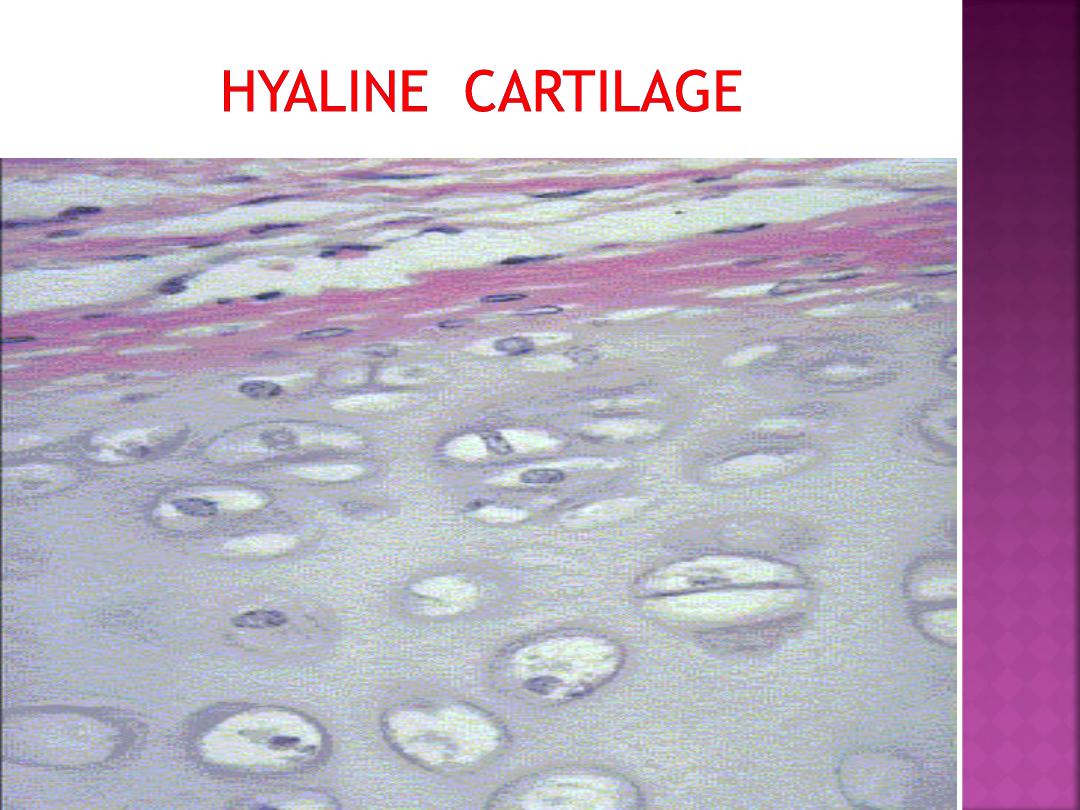
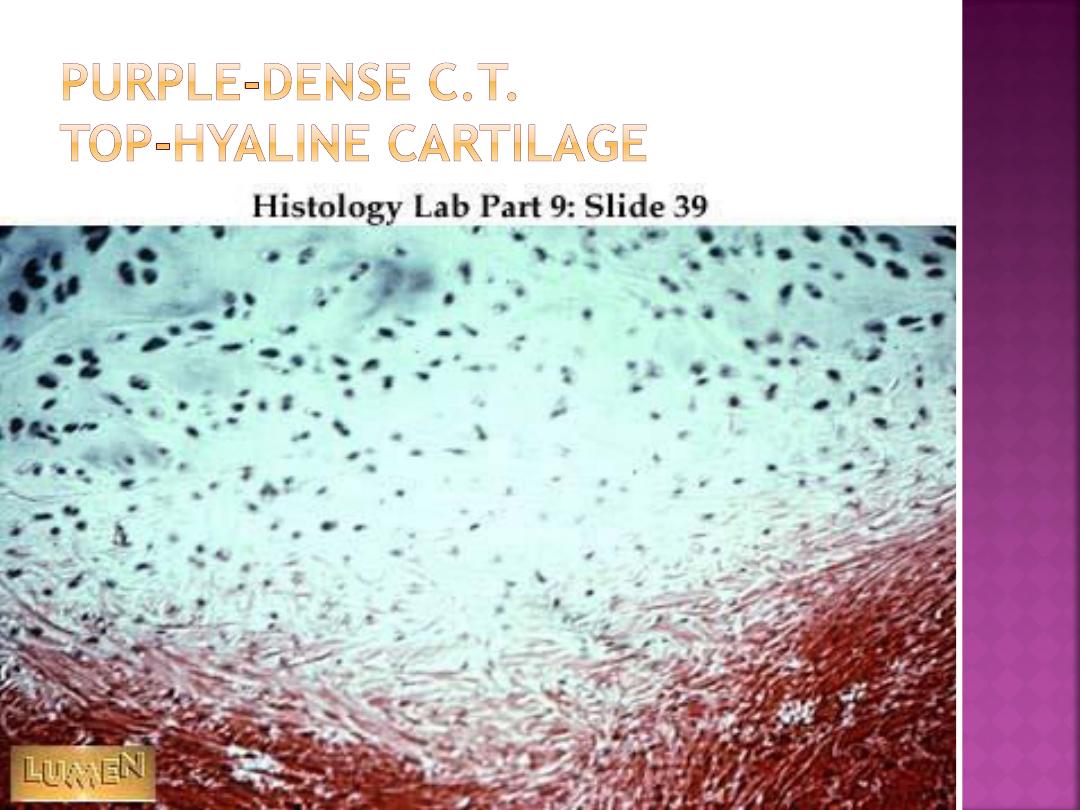
Hyaline cartilage is the most common type. In
embryos, hyaline cartilage serves as a skeletal
model for most bones. As the individual grows,
the cartilage bone model is gradually replaced
with bone by a process called
endochondral
ossification.
In adults, most of the hyaline
cartilage model has been replaced with bone,
except on the articular surfaces of bones, ends of
ribs (costal cartilage), nose, larynx, trachea, and
in bronchi. Here, the hyaline cartilage persists
throughout life and does not calcify.

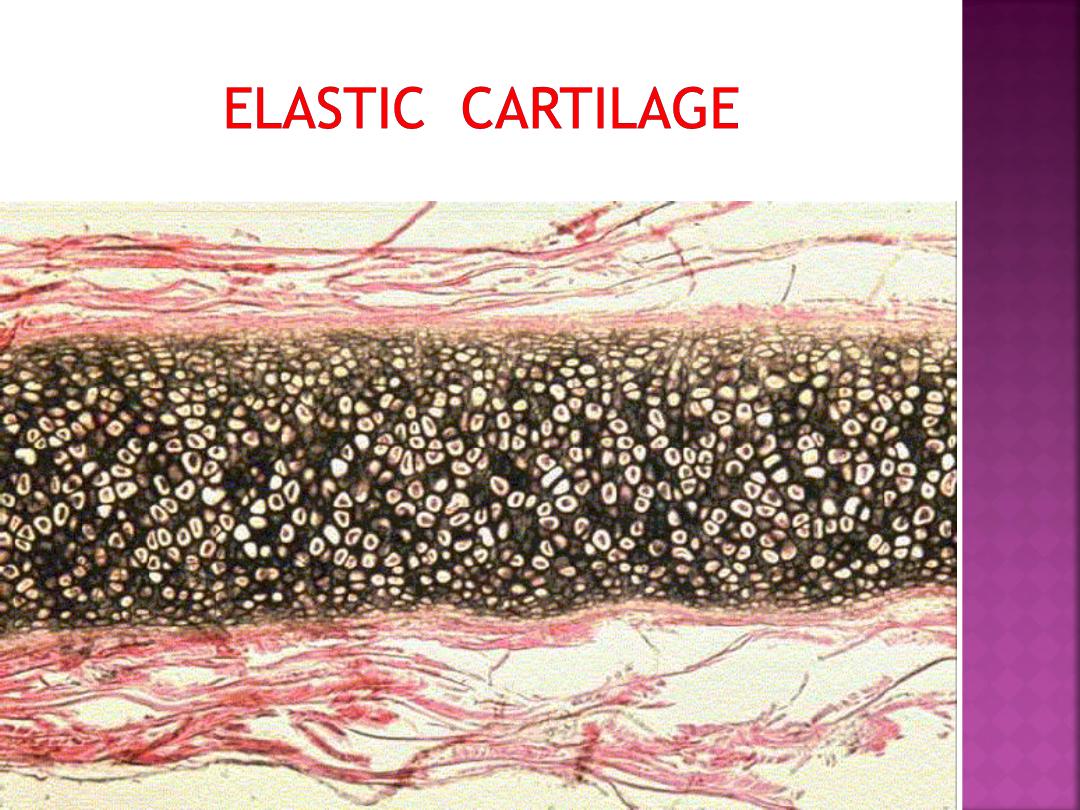
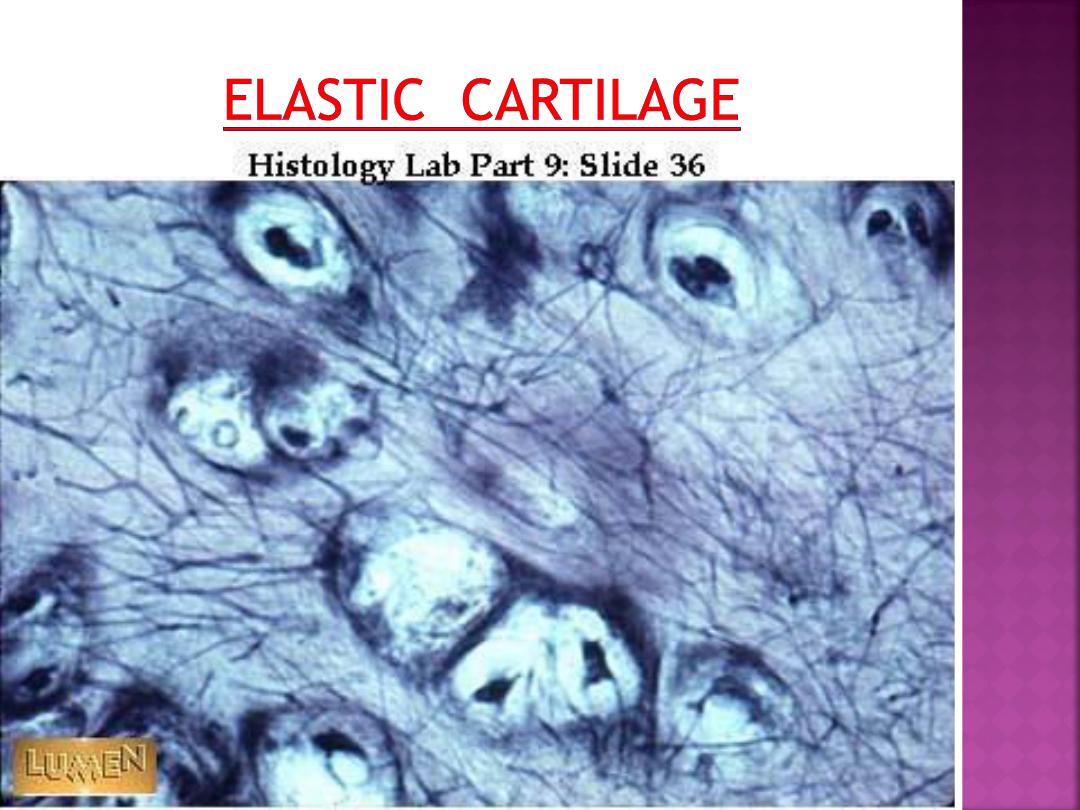
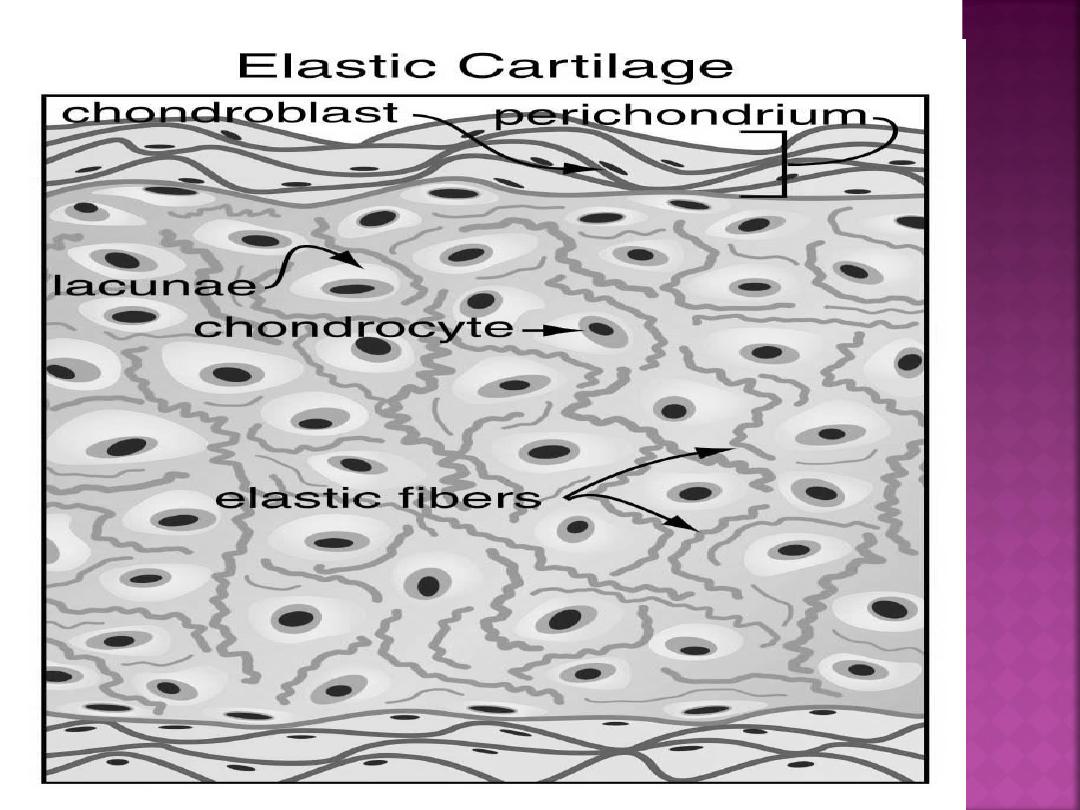
Elastic cartilage is similar in appearance to
hyaline cartilage, except for the presence
of numerous branching elastic fibers within
its matrix.
Elastic cartilage is highly flexible and
occurs in the external ear, walls of the
auditory tube, epiglottis, and larynx.

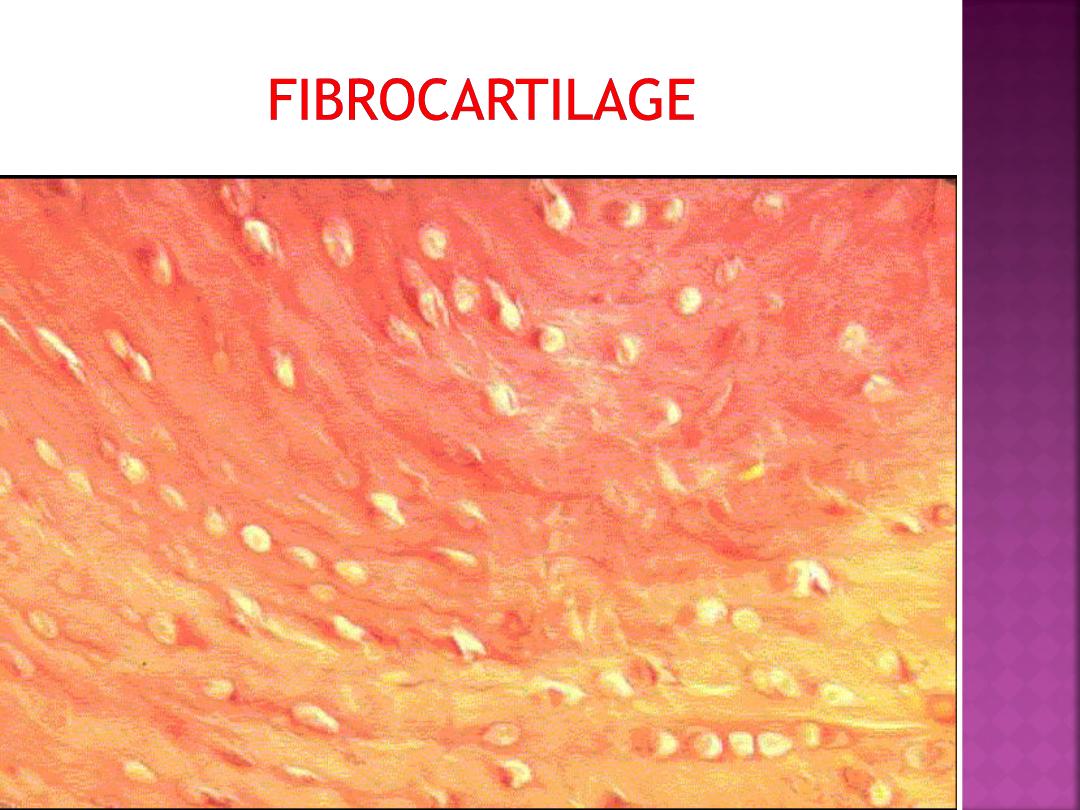
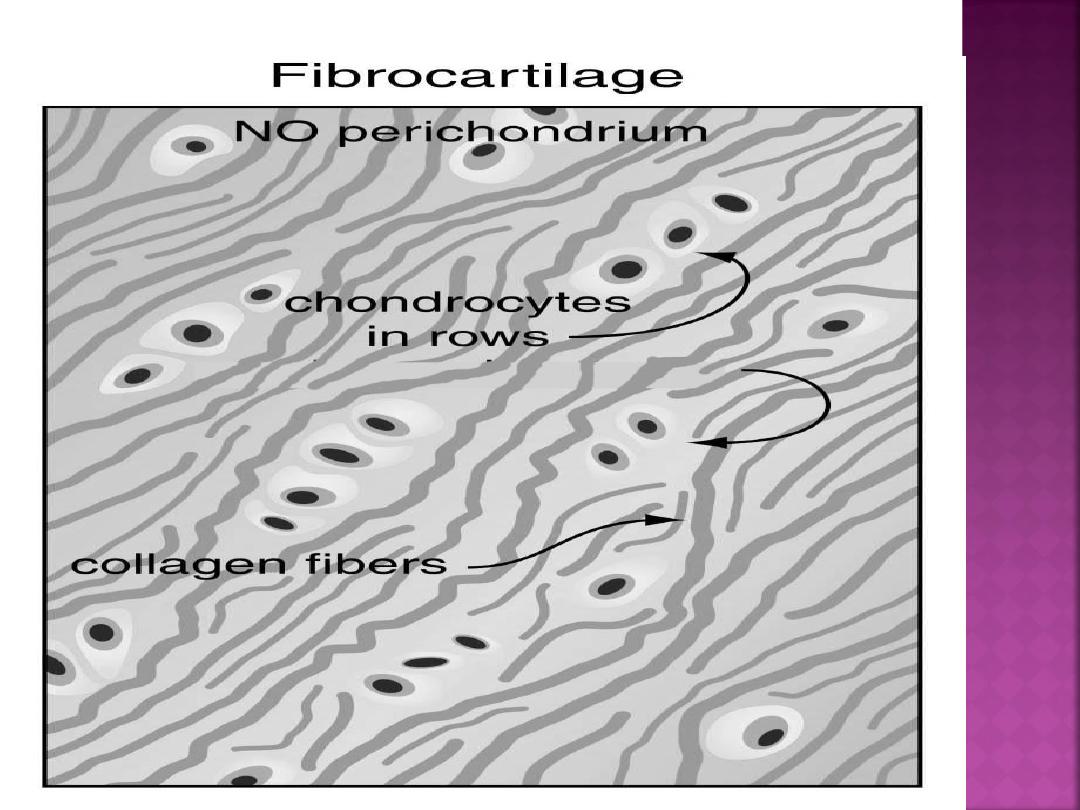
Fibrocartilage is characterized by large amounts
of irregular and dense bundles of coarse collagen
fibers in its matrix. In contrast to hyaline and
elastic cartilage, fibrocartilage consists of
alternating layers of cartilage matrix and thick
dense layers of type I collagen fibers.
Fibrocartilage has a limited distribution in
the body and is found in the intervertebral disks,
symphysis pubis, and certain joints.

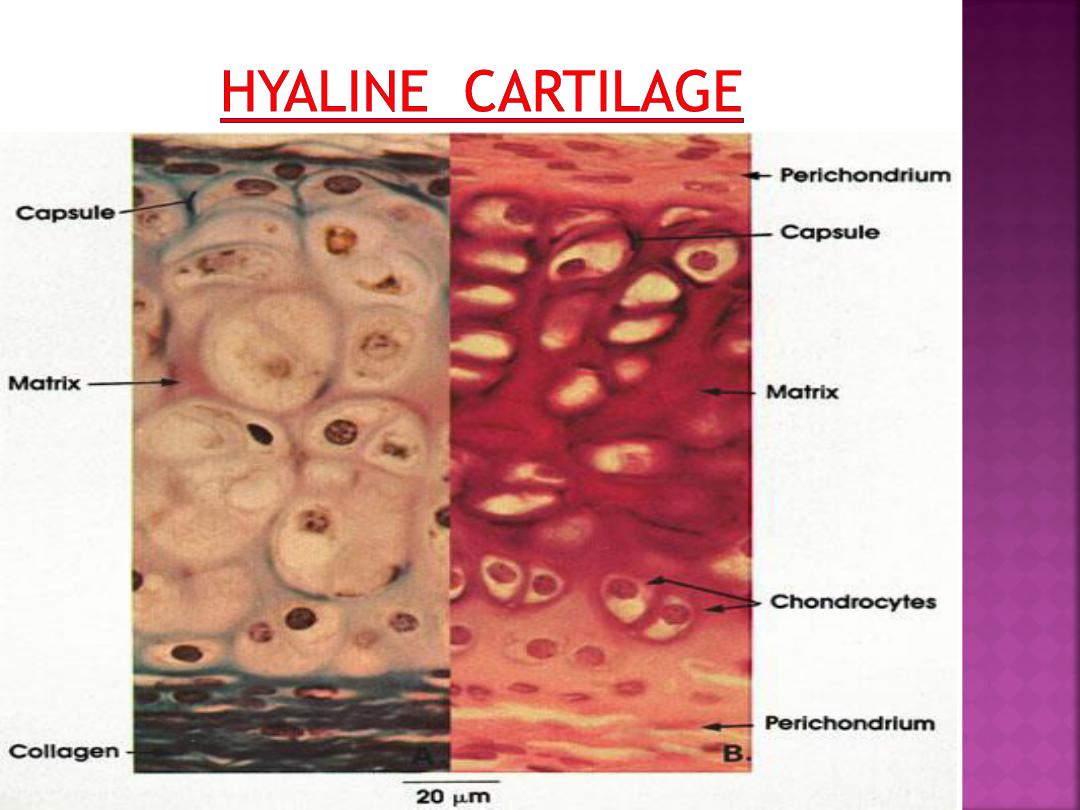
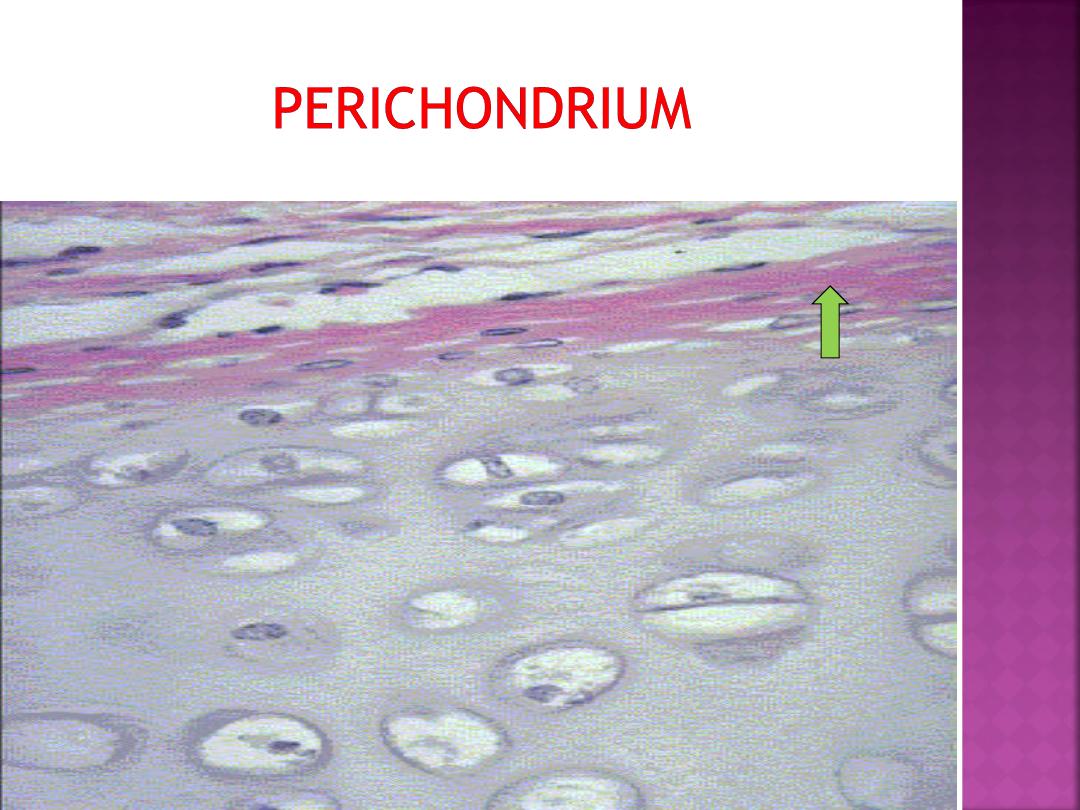
Most of the hyaline and elastic cartilage is
surrounded by a peripheral layer of vascularized,
dense, irregular connective tissue called the
perichondrium.
Its outer fibrous layer contains
type I collagen fibers and fibroblasts.
The inner layer of perichondrium is cellular
and chondrogenic.
Chondrogenic cells form the chondroblasts
that secrete the cartilage matrix.

the articulating surfaces of bones is not
lined by perichondrium. Similarly, because
fibrocartilage is always associated with dense
connective tissue fibers, it does not exhibit
an identifiable perichondrium.

Cartilage matrix is produced and maintained
by chondrocytes and chondroblasts.
The collagen or elastic fibers give cartilage
matrix its firmness and resilience.
The extracellular ground substance of
cartilage contains
sulfated glycosaminoglycans
and
hyaluronic acid
that are closely associated
with the elastic and collagen fibers within the
ground substance.

Also, cartilage matrix is highly hydrated
because of its high water content,which allows
for diffusion of molecules to and from the
chondrocytes.
Hyaline cartilage matrix consists of the fine type
II collagen fibrils embedded in a firm amorphous
hydrated matrix rich in proteoglycans and
structural glycoproteins.

In addition to type II collagen fibrils and
proteoglycans, cartilage matrix also contains
an adhesive glycoprotein called chondronectin.
These macromolecules bind to
glycosaminoglycans and collagen fibers,
providing adherence of chondroblasts and
chondrocytes to collagen fibers of surrounding
matrix.

Cartilage develops from primitive mesenchyme
cells that differentiate into
chondroblasts.
These cells divide mitotically and synthesize
the cartilage matrix and extracellular material.
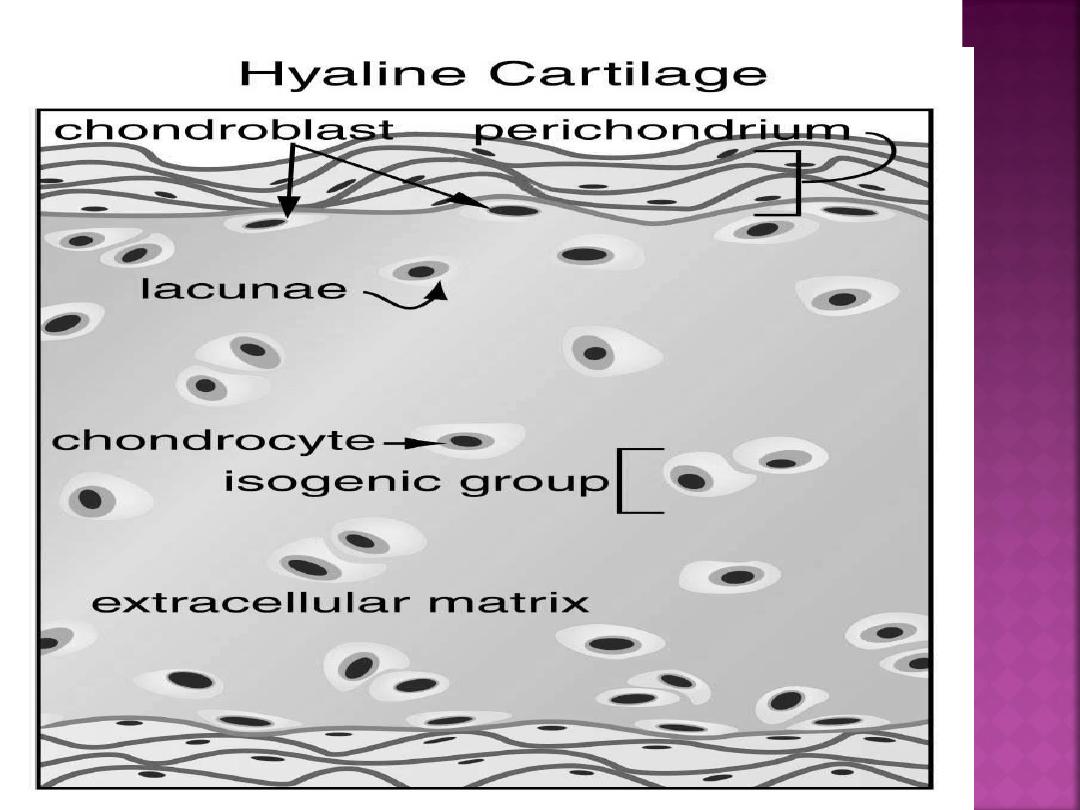
As the cartilage model grows, the individual
chondroblasts are surrounded by extracellular
matrix and become trapped in compartments
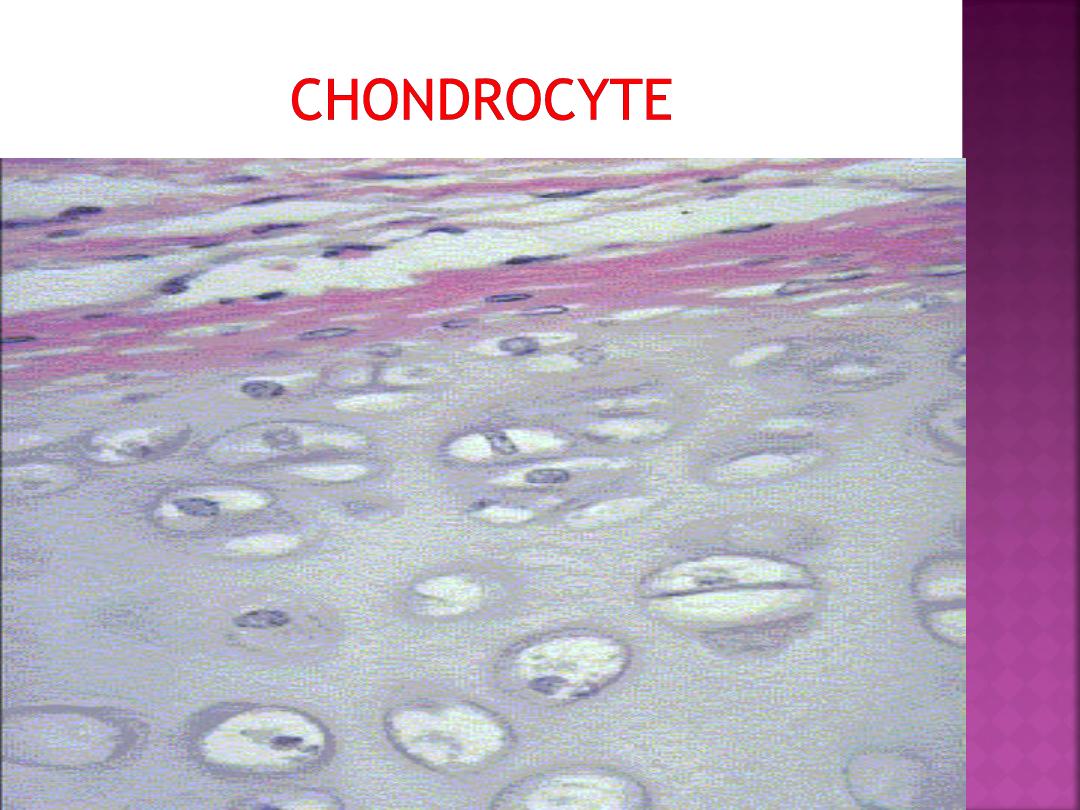
called lacunae (singular, lacuna).

In the lacunae are mature cartilage cells called
chondrocytes.
The main function of chondrocytes
is to maintain the cartilage matrix.
Some lacunae may contain more than one
chondrocyte; these groups of chondrocytes are
called
isogenous groups.
Mesenchyme cells can also differentiate into
fibroblasts that form the perichondrium, a
dense, irregular connective tissue layer that
invests the cartilage. The inner cellular layer of
perichondrium contains chondrogenic cells, which
can differentiate into
chondroblasts .

cartilage can simultaneously grow by two
different processes:
1- interstitial growth
2- appositional growth
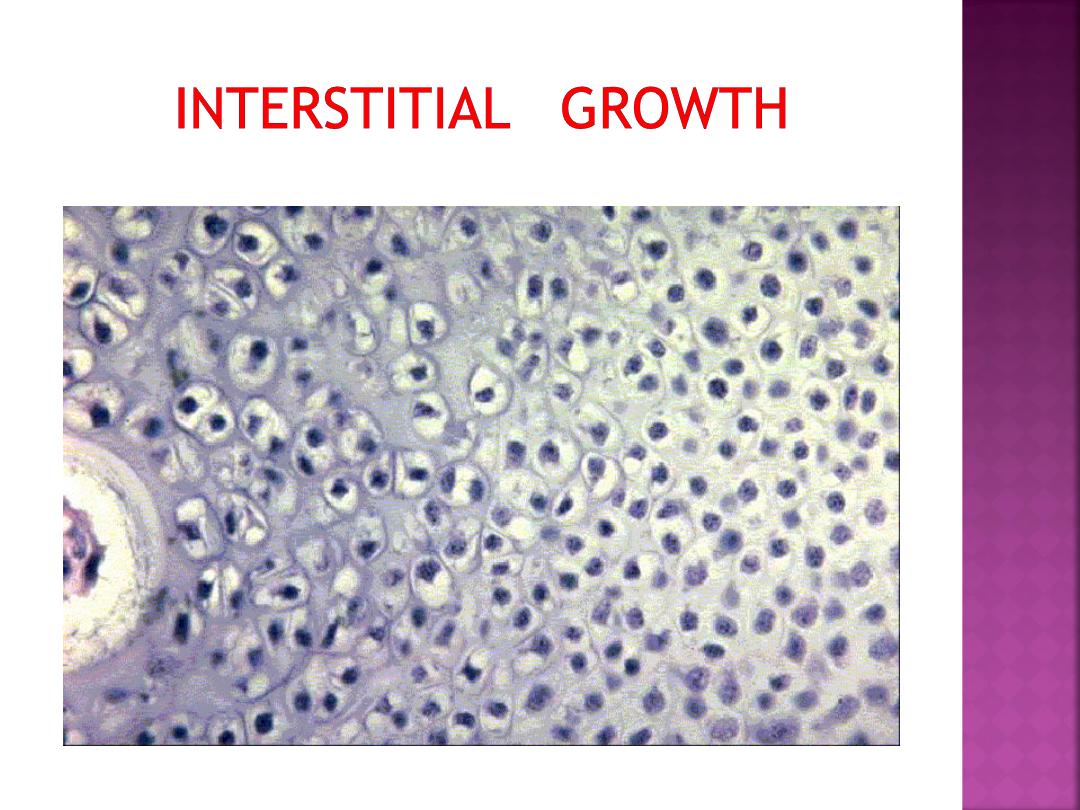
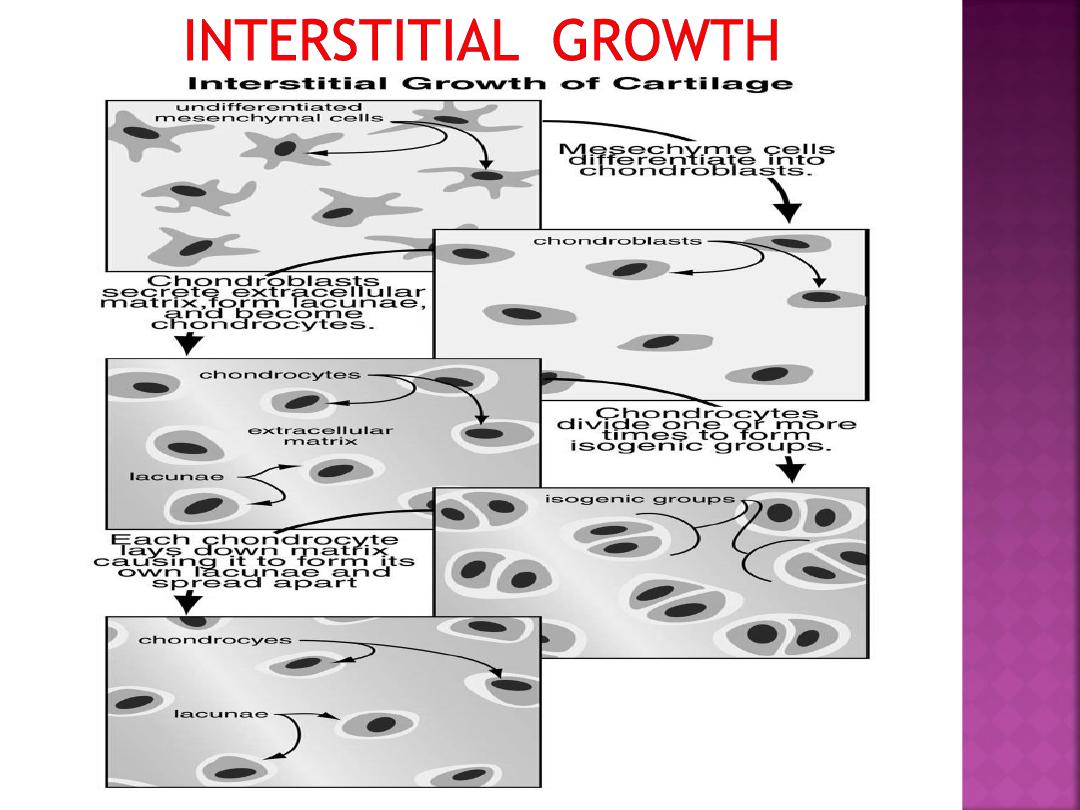

•As this occurs, the cartilage cells will undergo
divisions and continue to secrete matrix.
•Mesenchymal cells will aggregate and differentiate
into closely knit clusters of chondroblast
•These cells will begin to secrete collagen and
mucopolysaccharide matrix
.
•The matrix secretion will cause the chondroblasts
to be pushed apart
.
•

•In the case of hyaline cartilage, this
will result in some small clusters of
chondrocytes within the developing
matrix - isogenic groups.
Eventually the ground substance
becomes more rigid and the cartilage
cells (now chondrocytes) become
trapped in lacunae and can no longer be
pushed apart by secretion
.

This sort of growth of cartilage is
termed interstitial growth due to the
secretion of matrix into the interstitial
regions between cells or groups of cells.
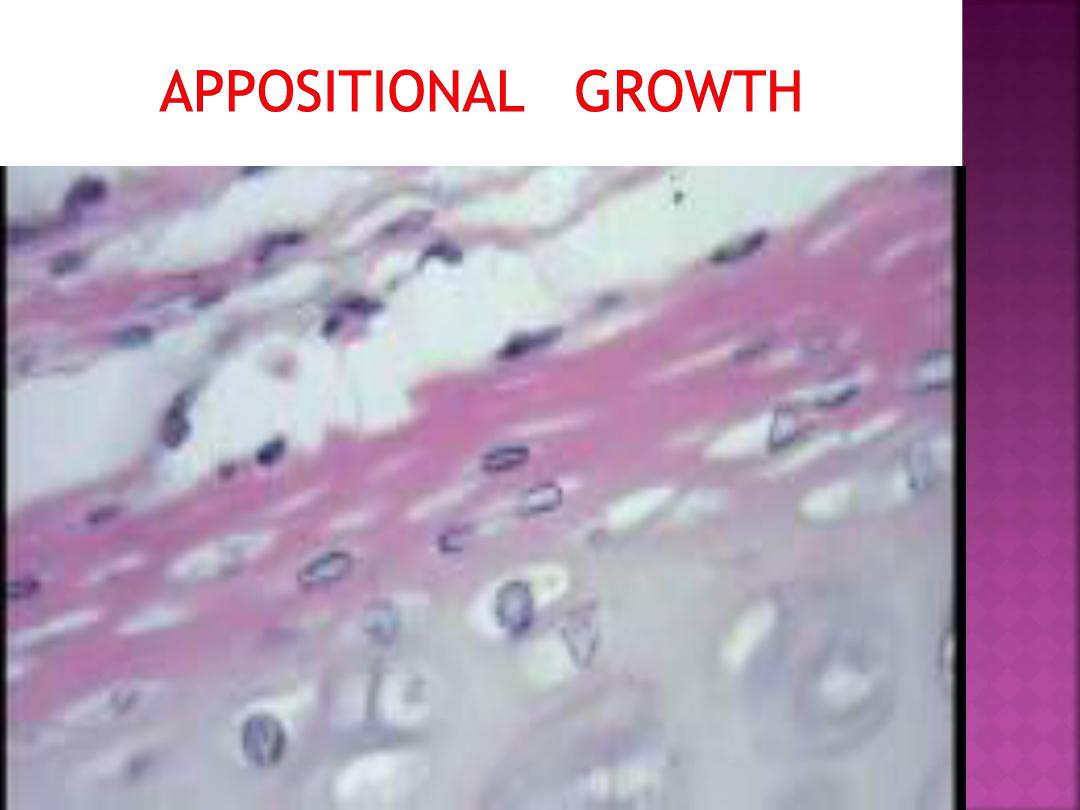

2. Appositional growth of cartilage
during embryogenesis and subsequent
juvenile development.
A layer of chondroblasts can lay
down matrix at the outer edge of a
mass of growing cartilage.
.
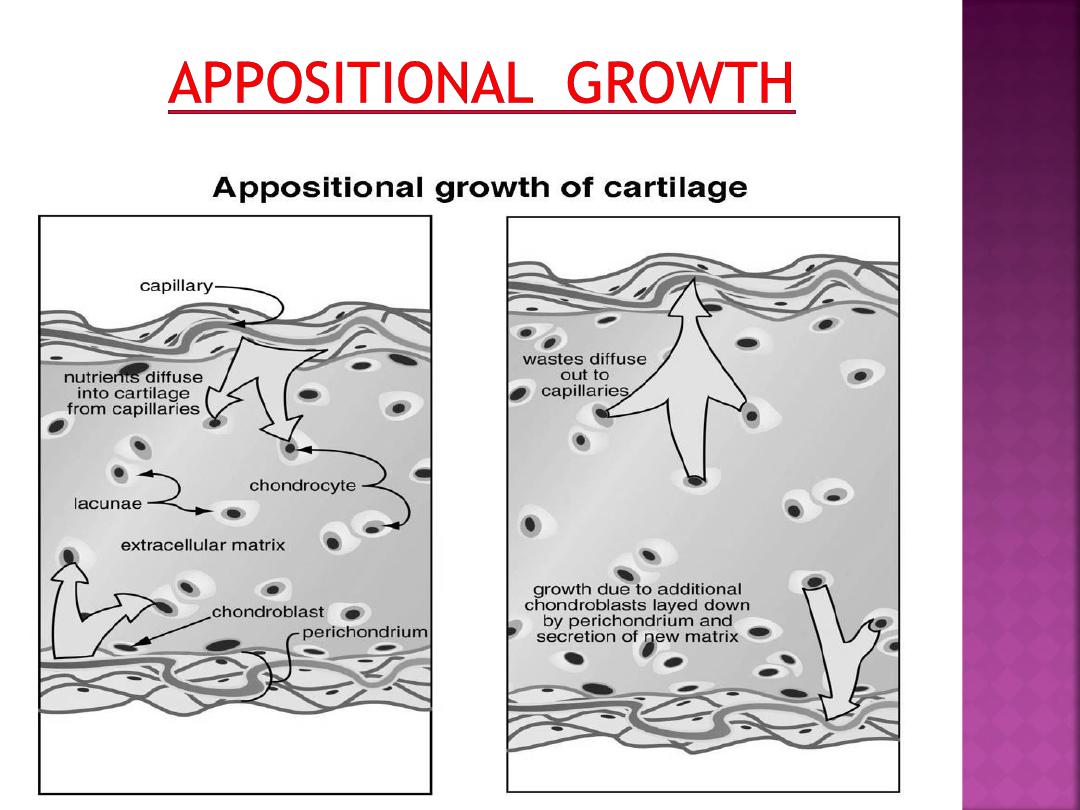

• Appositional growth can continue after
the perichondrim is formed. This is
accomplished by chondroblasts(and
perhaps fibroblasts) associated with the
perichondirum secreteing additional
ground substance.

Similar to cartilage, bone is also a special form of
connective tissue and consists of
cells, fibers
,
and
extracellular matrix
. Because of mineral
deposition in the matrix, bones become calcified.
As a result, bones , serve as a rigid skeleton for
the body, and provide attachment sites for
muscles and organs.
Bone also protects the brain in the skull, heart
and lungs in the thorax, and urinary and
reproductive organs between the pelvic bones.
In addition, bones function in hemopoiesis (blood
cell formation).

Two types of bone :
1-compact bone
2- cancellous bone (spongy)or trabecular
In long bones, the outer cylindrical part is the
dense compact bone.
The inner surface of compact bone adjacent to
the marrow cavity is the cancellous (spongy)
bone.
Cancellous bone contains numerous
interconnecting areas and is not dense;
however, both types of bone have the same
microscopic appearance.
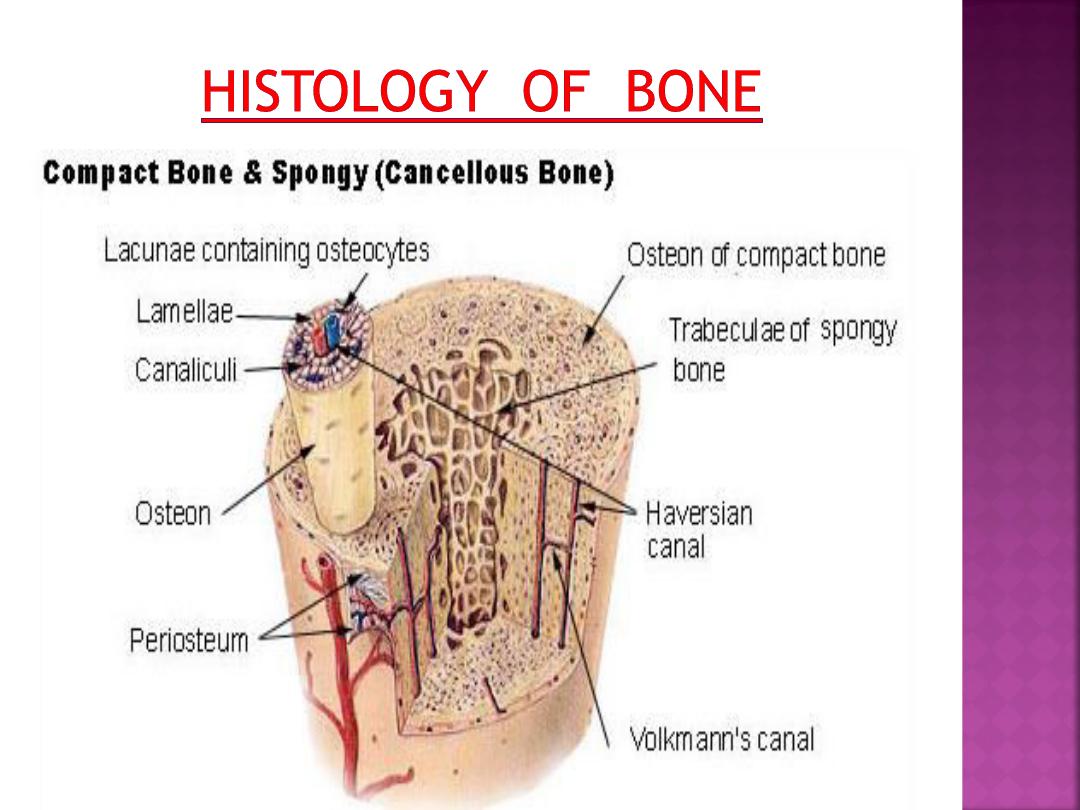

In compact bone, the collagen fibers are
arranged in thin layers of bone called
lamellae
that are parallel to each other in the periphery
of the bone, or concentrically arranged around a
blood vessel.
In a long bone, the outer circumferential
lamellae are deep to the periosteum. Inner
circumferential lamellae surround the bone
marrow cavity.

Concentric lamellae surround the canals
with blood vessels, nerves, and loose connective
tissue called the
osteons (Haversian systems).
The space in the osteon that contains
blood vessels and nerves is the central
(Haversian) canal.Most of the compact bone
consists of osteons. Lacunae with osteocytes and
connected via canaliculi are found between the
lamellae in each osteon .


A second system of canals, called
Volkmann's canals, penetrates the bone more or
less perpendicular to its surface. These canals
establish connections of the Haversian canals
with the inner and outer surfaces of the bone.
Vessels in Volkmann's canals communicate with
vessels in the Haversian canals on the one hand
and vessels in the endosteum on the other.
A few communications also exist with vessels in
the periosteum.

The matrix of trabecular bone is also deposited
in the form of lamellae. In mature bones,
trabecular bone will also be lamellar bone.
However, lamellae in trabecular bone do not
form Haversian systems. Lamellae of trabecular
bone are deposited on preexisting trabeculae
depending on the local demands on bone
rigidity.
Osteocytes, lacunae and canaliculi in
trabecular bone resemble those in compact
bone.
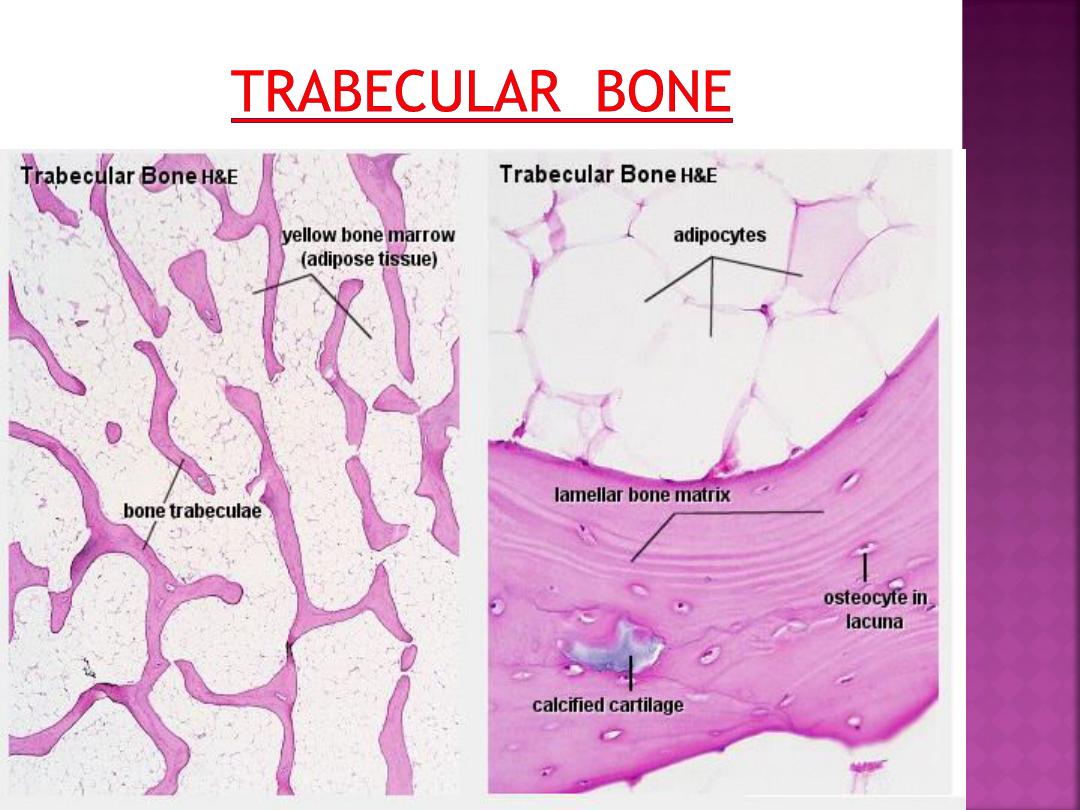

Bone matrix consists of collagen fibres (about
90% of the organic substance) and ground
substance.
Collagen type I is the dominant collagen form
in bone. The hardness of the matrix is due to
its content of inorganic salts (hydroxyapatite;
about 75% of the dry weight of bone), which
become deposited between collagen fibres.

Calcification begins a few days after
the deposition of organic bone substance (or
osteoid) by the osteoblasts. Osteoblasts are
capable of producing high local concentration
of calcium phosphate in the extracellular
space, which precipitates on the collagen
molecules. About 75% of the hydroxyapatite
is deposited in the first few days of the
process, but complete calcification may take
several months.

Developing and adult bones contain
four different cell types:
osteoprogenitor cells,
osteoblasts,
osteocytes,
osteoclasts.

are undifferentiated, pluripotential stem
cells derived from the mesenchyme. These cells
are located on the inner layer of connective tissue
periosteum and in the single layer of internal
endosteum that lines the marrow cavities,
osteons (Haversian system) .
During bone development, osteoprogenitor
cells proliferate by mitosis and differentiate into
osteoblasts, which then secrete collagen fibers
and the bony matrix.

Osteoblasts (or bone forming cells).
may form a low columnar "epitheloid
layer" at sites of bone deposition. They
contain plenty of rough endoplasmatic
reticulum (collagen synthesis) and a large
Golgi apparatus. As they become trapped
in the forming bone they differentiate into
osteocytes.
Osteocytes.
Osteocytes contain less endoplasmatic
reticulum and are somewhat smaller than
osteoblasts.

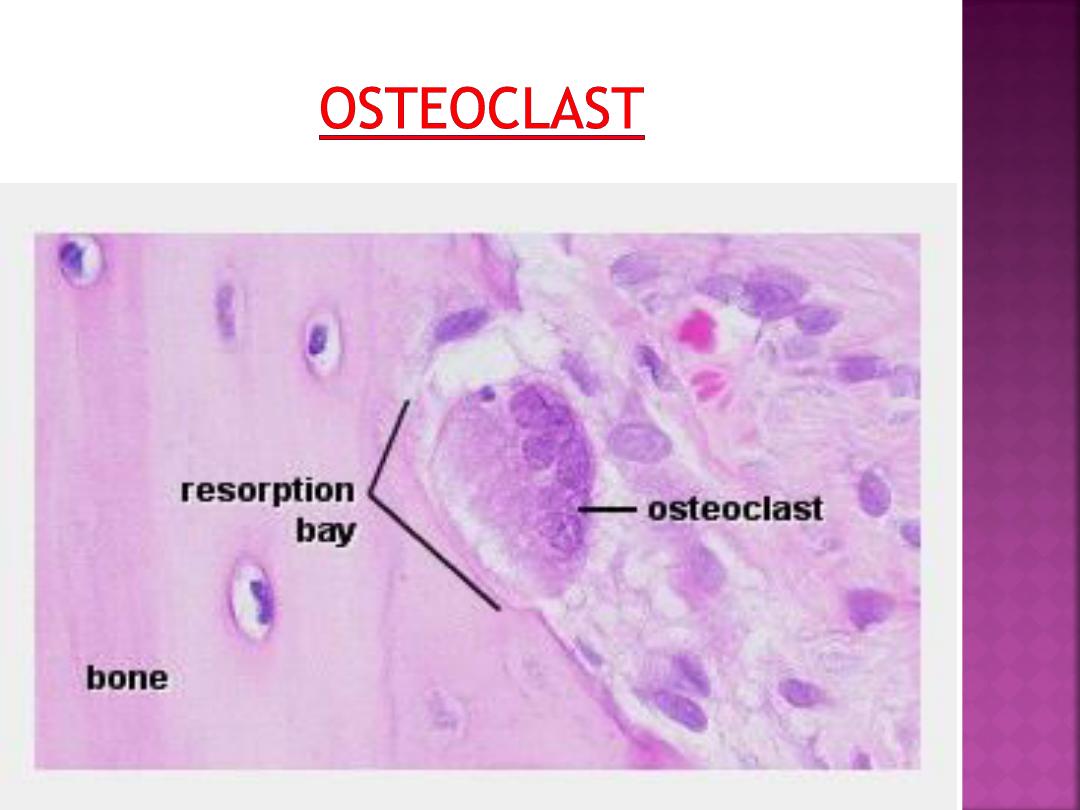
are very large , multi-nucleated (about 5-
10 visible in a histological section, but up
to 50 in the actual cell) bone-resorbing
cells.
They arise by the fusion of monocytes
(macrophage precursors in the blood) or
macrophages.
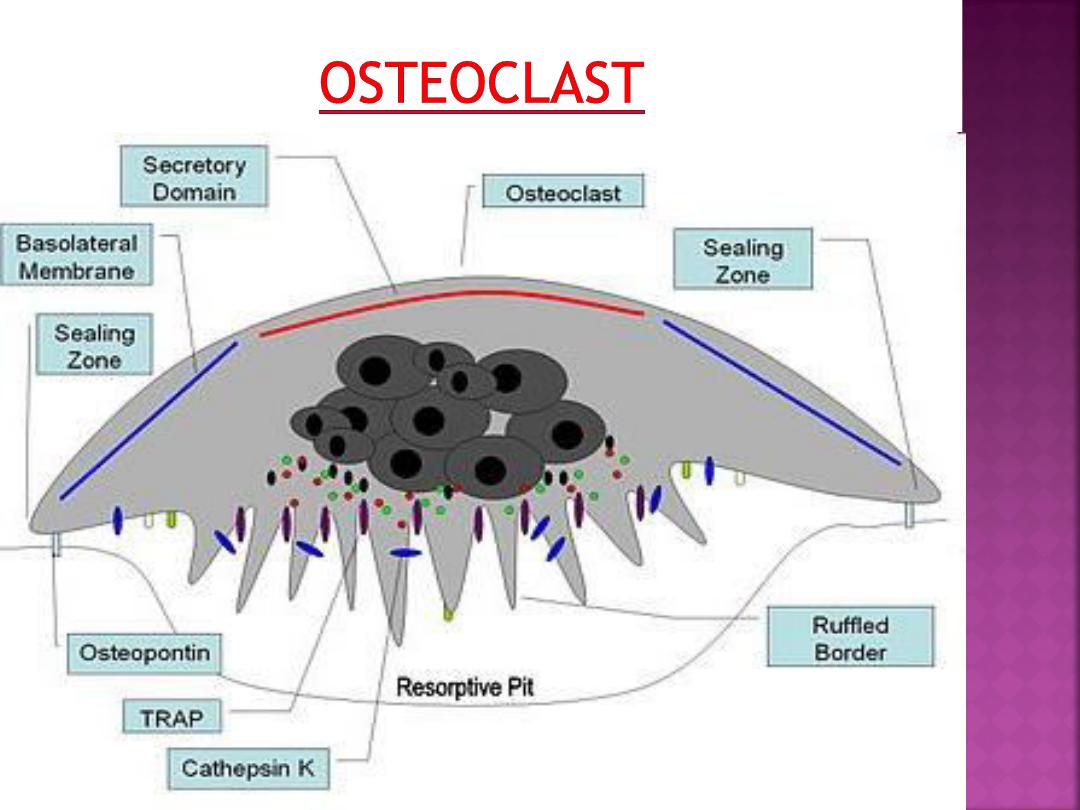
Osteoclasts attach themselves to the
bone matrix and form a tight seal at the rim
of the attachment site. The cell membrane
opposite the matrix has deep invaginations
forming a
ruffled border
.

They released enzymes break down the
collagen fibres of the matrix.
Osteoclasts are stimulated by parathyroid
hormone (produced by the parathyroid gland)
and inhibited by calcitonin (produced by
specialised cells of the thyroid gland).
Osteoclasts are often seen within the
indentations of the bone matrix that are
formed by their activity (resorption bays or
Howship's lacunae).

Modeling is a process in which bone is sculpted
during growth to ultimately achieve its proper
shape. Modeling is responsible for the
circumferential growth of the bone and
expansion of the marrow cavity, and
enlargement of the cranial vault curvature.
Remodeling is a continuous process throughout
life, in which damaged bone is repaired, ion
homeostasis is maintained, and bone is
reinforced for increased stress. In adults, the
remodeling rate varies in different types of
bones.

Bone formation or development occurs by
these mechanisms:
1-endochondral ossification
2- intramembranous ossification

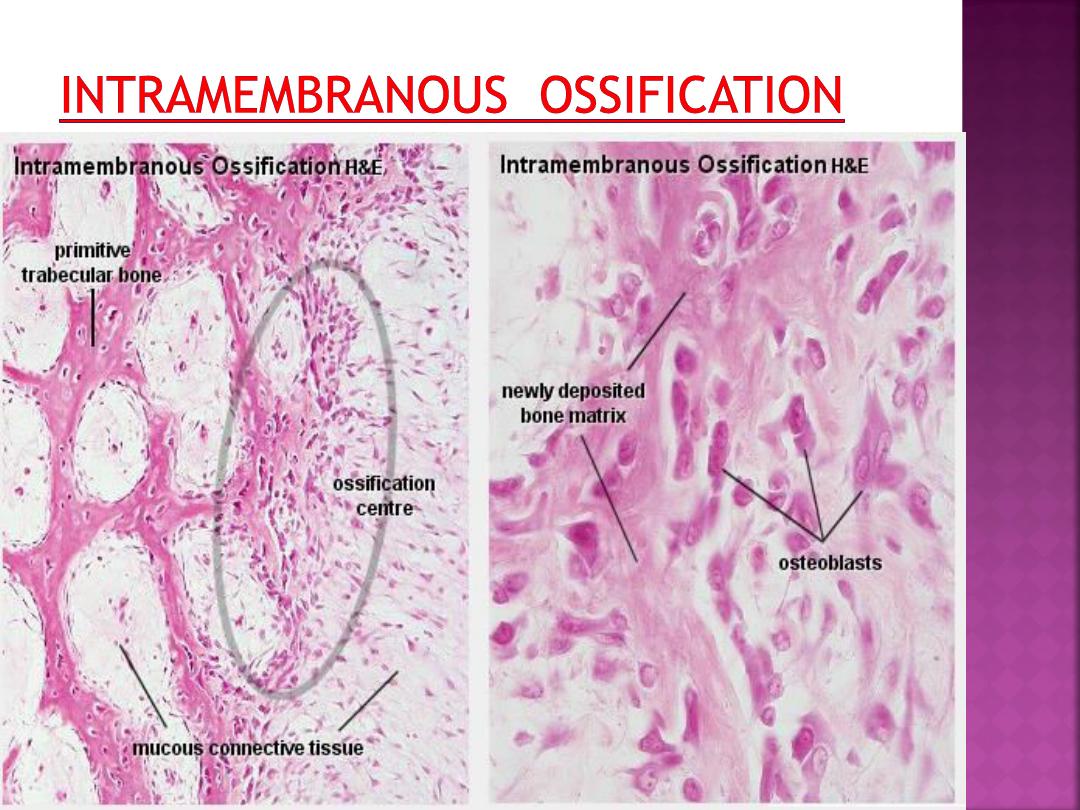
Intramembranous ossification occurs within a
membranous, condensed plate of
mesenchymal cells.
At the initial site of ossification (ossification
centre) mesenchymal cells (osteoprogenitor
cells) differentiate into osteoblasts.
The osteoblasts begin to deposit the organic
bone matrix, the osteoid. The matrix
separates osteoblasts, which, from now on,
are located in lacunae within the matrix

The collagen fibres of the osteoid form a
woven network without a preferred
orientation, and lamellae are not present at
this stage.
Because of the lack of a preferred orientation
of the collagen fibres in the matrix, this type
of bone is also called woven bone. The
osteoid calcifies leading to the formation of
primitive trabecular bone.
Further deposition and calcification of
osteoid at sites where compact bone is
needed leads to the formation of primitive
compact bone.

•
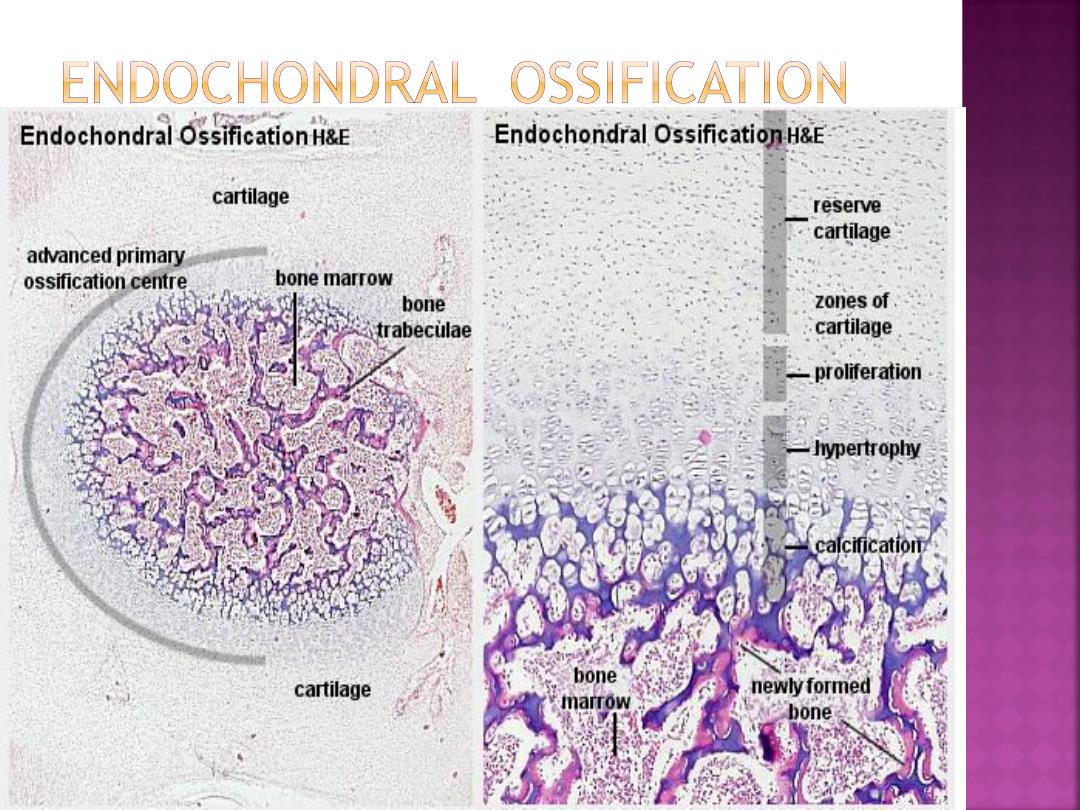
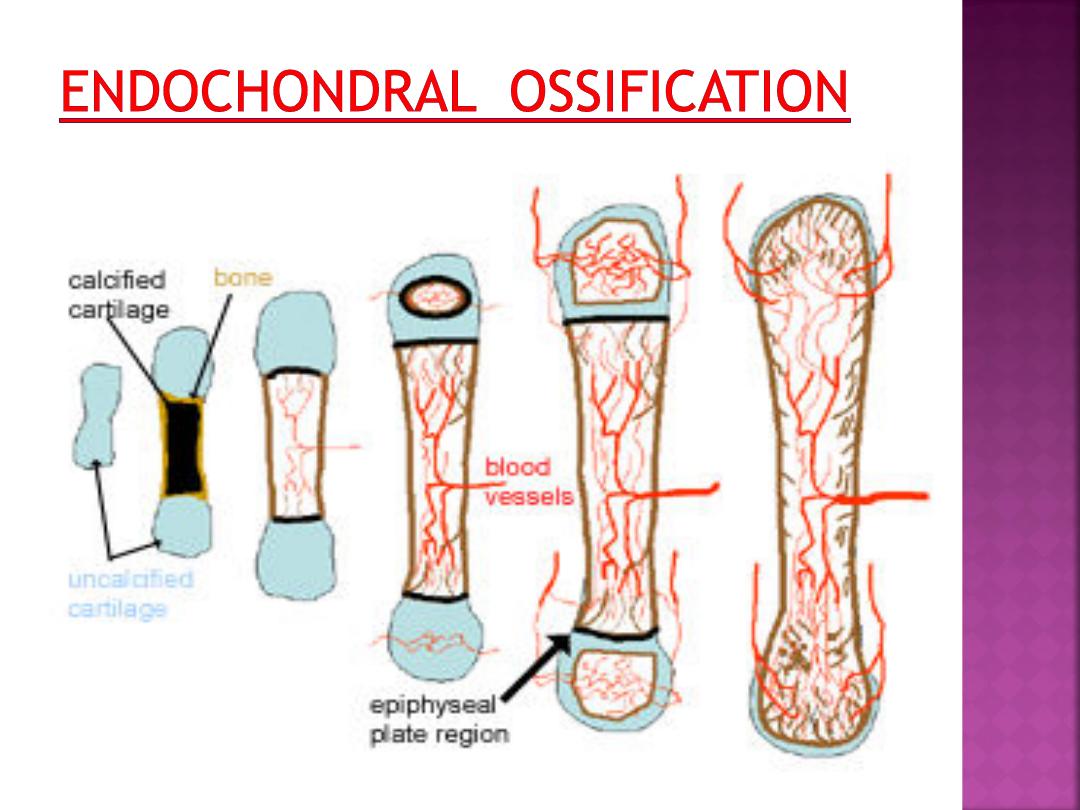
The bone is formed onto a temporary cartilage
model.
•
The cartilage model grows (zone of
proliferation), then chondrocytes mature (zone
of maturation) and hypertrophy (zone of
hypertrophy), and growing cartilage model starts
to calcify.
•
As this happens, the chondrocytes are far from
blood vessels, and are less able to gain nutrients
etc, and the chondrocytes start to die (zone of
cartilage degeneration). The fragmented
calcified matrix left behind acts as structural
framework for bony material.

Osteoprogenitor cells and blood vessels
from periosteum invade this area, proliferate and
differentiate into osteoblasts, which start to lay
down bone matrix (osteogenic zone).
•
In the fetus, the primary ossification centre
forms first in the diaphysis. Later on a secondary
ossification centre forms in the epiphysis.
•
Cartilage is replaced by bone in the epiphysis and
diaphysis, except in the epiphyseal plate region.
Here the bone continues to grow, until maturity
(around 18 years old).

Primary and secondary ossification centres do
not merge before adulthood.
Between the diaphysis and the epiphyses a
thin sheet of cartilage, the epiphyseal plate,
is maintained until adulthood.
By continuing cartilage production, the
epiphyseal plate provides the basis for rapid
growth in the length of the bone.
Cartilage production gradually ceases in the
epiphyseal plate as maturity is approached.
The epiphyseal plate is finally removed by the
continued production of bone from the
diaphyseal side.

As the zone of ossification moves in the
direction of the future epiphyses. This
process creates the marrow cavity of
the bones.
Simultaneously, bone is removed from
the endosteal surface and deposited on
the periosteal surface of the compact
bone which forms the diaphysis.
This results in a growth of the diameter
of the bone .