
Unit 4: Virology
251
Lecture 1+2+3 - Introduction
Virus (Virion)
It is the smallest infectious agents and contains one kind
of nucleic acid as their genome (DNA or RNA). Size
range from (20nm-300nm in diameter). Viruses replicates
only inside the living cells of other organisms and can
infect all types of life forms, from animals and plants to
bacteria and archaea
I-Viral mucleic acid
The viral nucleic acid is located internally and can be
either double or single stranded DNA or RNA.
The nucleic acid is either circular or linear.
The DNA is always a single molecule (double or single);
RNA can exist either as a single molecule or in several
pieces.
Almost all viruses contain only a single copy of their
genome (haploid) but retroviruses are the exception (RNA
genome of two copies (Diploid)).
II-Viral capsid
The capsid is the “shell” of virus-coded protein that
encloses the nucleic acid and is more or less closely
associated with it. The combination of these two
components (Capsid and NA) is often termed the
nucleocapsid, especially if they are closely associated as
in the myxoviruses.
The capsid is made up of subunits, the capsomers, the
number of which varies but is specific and constant for
each viral species. These (capsomeres) are spherical or
cylindrical structures composed of several polypeptides.
The capsid protects the nucleic acid from degradation. In
all except enveloped viruses, it is responsible for the
attachment of the viruses to the host cell.
Viral Symmetry
The arrangement of capsomers gives the virus structure
and its geometric symmetry. Viral nucleocapsids have
two forms of symmetry:
1) Icosahedral:
(polyhedrons with 20 equilateral triangular
faces)
in which the capsomers are arrangred in 20
triangles that form a symmetric figure with approximate
outline of the sphere.
2) Helical: in which the capsomers are arranged in a hollow
coil that that appears rod-shaped.
All human viruses that have a helical nucleocapsid are
enclosed by an outer membrane called an envelope while
icosaheral nucleocapsid can be either enveloped or non-
enveloped (naked).
3) Complex symmetry. Complex structural patterns are
found in bacteriophages and the smallpox virus.
III- Viral proteins
Structural proteins of viruses have several important
functions:
1) Facilitate transfer of the viral nucleic acid from one host
cell to another.
2) Protect viral genome against inactivation by nucleases
3) Participate in the attachment of the virus particle to
susceptible cell.
4) Provide the structural symmetry of the virus particle.
5) Determine the antigenic characteristics of the virus.
There are frequently glycoproteins in the form of spike
like projections on the surface, which attach to host cell
receptors during the entry of the virus into the cell.
Another protein is the Matrix protein mediates the
interaction between the capsid protein and the envelope.
Non-Structural proteins
Some viruses carry enzymes inside the virions which are
essential for the initiation of the viral replicative cycle
when the virus enter host cell e.g.: a) RNA polymerase
which is carried by negative sense RNA virus that is
needed to copy the first mRNA. b) reverse transcriptase in
retroviruses that makes a DNA copy of the viral RNA.
Some viruses contain regulatory proteins in the virion in a
structure called the tegument which is located between
nucleocapsid and the envelope. These regulatory proteins
include transcription and translation factors that control
either viral or cellular processes. Herpes simplex and
cytomegalovirus have well characteristics tegument.

Unit 4: Virology
251
IV-Viral envelope
It is a lipoprotein membrane composed of lipid derived
from the host cell membrane and protein that is virus
specific.
Enveloped viruses are more sensitive to heat, ether,
detergents, and dryness. They are transmitted by direct
contact.
Non-enveloped virus (naked) are more stable and
transmitted by indirect contact.
Classification of viruses
The taxonomic system used for viruses is artificial (i.e., it
does not reflect virus evolution) and is based on the
following morphological and biochemical criteria:
1) Genome: DNA or RNA genome (important basic
differentiation of virus types!) as well as configuration of
nucleic acid structure: single-stranded (ss) or double-
stranded (ds); RNA viruses are further subclassified
according to plus and minus polarity.
2) Capsid symmetry: cubic, helical, or complex symmetry.
3) Presence or absence of an envelope.
4) Diameter of the virion, or of the nucleocapsid with
helical symmetry.
Atypical virus –like agents
1) Defective viruses: are viruses composed of viral N.A and
proteins but cannot replicate without a "helper" virus,
which provides the missing function. There is usually a
mutation or deletion of a part of their genetic material
2) Pseudovirions: contain host cell DNA instead of viral
DNA within the capsid.they are formed during infection
with certain viruses when host cell DNA is fragmented
and pieces of it are incorporated within the capsid protein.
3) Viriods: single molecule of circular RNA without a
protein coat or envelope.
4) Prions: are infectious particles that are composed solely
of protein. They are implicated as the cause of certain
slow disease which called transmissible spongiform
encephalopathies.
Viral replication
The growth curve in figure 1 shows that first event is
disappearance of the virus as represented by the solid line
dropping to the x axis. Although the virus particle is no
longer present, the viral nucleic acid continues to function
and begins to accumulate within the cell. The time during
which no virus is found inside the cell is known as the
Eclipse period. It ends with the appearance of the virus
(solid line). The latent period is defined as the time from
the onset of infection to the appearance of the virus
extracellularly. At the end of latent period, alterations of
cell morophology accompanied by marked derangement
of cell function; this is called cytopathic effect (CPE)
which culminates in the lysis and death of cells. These
CPE can be seen under the light microscope.
Figure 1: Viral growth curve.
Specific events during the growth cycle
I-Early events ( attachment, penetration,
&uncoating)
The proteins on the surface of the virion attach to specific
receptor proteins on the cell surface through weak non-
covalent bonding. The specifity of the attachment
determines the host range of the virus.
The virus particle penetrates by being engulfed in a
pinocytic vesicle, within which the process of uncoating
begins.
The receptors for viruses on the cell surface are proteins
that have other functions in the life of the cell.
NOTES:
1- Infectious nucleic acid is purified viral RNA or DNA
(without any protein) that can carry out the entire viral
growth cycle & result in the production of complete virus
particle.
2- All viruses are "infectious" but not all purified genomes
are infectious.
II- Middle events (Gene Expression and Genome
Replication)
The first step in viral gene expression is mRNA synthesis.
It is at this point that viruses follow different pathways
depending on the nature of their nucleic acid and the part
of the cell in which they replicate.

Unit 4: Virology
251
DNA viruses: Most have both positive and negative
strand (Ds) (except parvovirus has ssDNA), but only +
strand is read.
All DNA Viruses replicate in the nucleus except
poxvirus which has its own RNA polymerase to replicate
in the cytosol
Retrovirus: Carries reverse transcriptase, which
converts RNA to DNA in the nucleus, inserting DNA
copy of genome into host cell DNA then using host
transcription to mRNA.
Positive ssRNA viruses (e.g. poliovirus): Use RNA
directly as mRNA to begin translation immediately in the
cytosol
Negative ssRNA viruses: Must be transcribed to +ssRNA
before being translated. Uses own RNA-dependent RNA
polymerase to do transcription in the cytosol (except
Influenza in nucleus).
Single RNA and segmented RNA are both in this
category, though segmented RNA viruses may be
ambisense (some -, some +ssRNA)
Double-stranded RNA: Carries its own RNA-dependent
RNA polymerase to translate into mRNA in cytosol (+
strand hydrogen bonded to - strand so cannot be used as
mRNA). Reoviruses
Early proteins are
enzymes
required for replication of
viral genome (e.g. Hepadnavirsus produce reverse
transcriptase to regenerate viral DNA from RNA
intermediate).
Most viruses make a virus-encoded polymerase
(
replicase )to make copies of genome, although some use
host cell polymerases for this
Late proteins
: structural proteins for progeny viruses,
often including precursor polypeptides and virus-coded
proteases to cleave these polypeptides into final
capsomer products (e.g. picornaviruses have single
polypeptide that has intrinsic protease, retroviruses)
III- Late events (Assembly and Release)
The progeny particles are assembled by packing the viral
nucleic acid within the capsid proteins.
Un enveloped viruses are released by cell rupture.
Enveloped viruses are released by budding through the
outer cell membrane Except Herpes virus by budding
from the nuclear membrane.
Lysogeny
Is the process by which viral DNA becomes integrated
into host cell DNA, replication stops and no progeny virus
is made. Later if DNA is damaged by for example by UV
light, viral DNA is excised from the host cell DNA , and
progeny viruses are made. The integrated viral DNA is
called a Prophage. Bacterial cells carrying a prophage
can acquire a new trait, such as the ability to produce
exotoxins such as Diphtheria toxin.
Lysogenic conversion is the term used to indicate that the
cell has acquired a new trait as a result of the integrated
prophage. Lysogenic conversion is mediated by
transduction ( transfer of gene from one bacterium to
another by viruses).
Relationship of lysogeny in bacteria to latency in
human
Members of the Herpesvirus family, such as herpes
simplex virus (HSV), Varicella zoster virus (VZV),
Cytomegalovirus (CMV), and Epstein-Barr virus (EBV),
exhibit a latency – the phenomenon in which no or very
little virus is produced after the initial infection but at
some later time, reactivation and full virus replication
occur.
How Herpesvirus initiate and maintain the latent state?
Shortly after the virus infects neurons, a set of "Latency-
associated transcripts" (LATS) are synthesized. These
are non – coding, regulatory RNAs that suppress viral
replication. The precise mechanism by which they do so
is unclear. Reactivation of viral replication at a later time
occurs when the genes encoding LATS are excised.
CMV employs different mechanisms. The CMV genome
encode microRNAs that inhibit the translation of mRNAs
required for viral replication. Also, the CMV genome
encodes both a protein and an RNA that inhibits
apoptosis. This allows the infected cell to survive.
Virus genetics
Mutation
Mutations are changes in the base sequence of a nucleic
acid (base substitution, deletion and frame shift), resulting
in a more or less radical alteration of the resulting protein.
Medically important are mutant virus with weakened
virulence that have retained their antigenicity and
replication capabilities intact. These are known as
“attenuated” viruses. They are the raw material of live
vaccines.
Interaction between viruses
When two genetically distinct viruses infect a cell, three
different phenomena can ensue:
1) Recombination
Is the exchange of genes between two chromosomes that
is based on crossing over within regions of significant
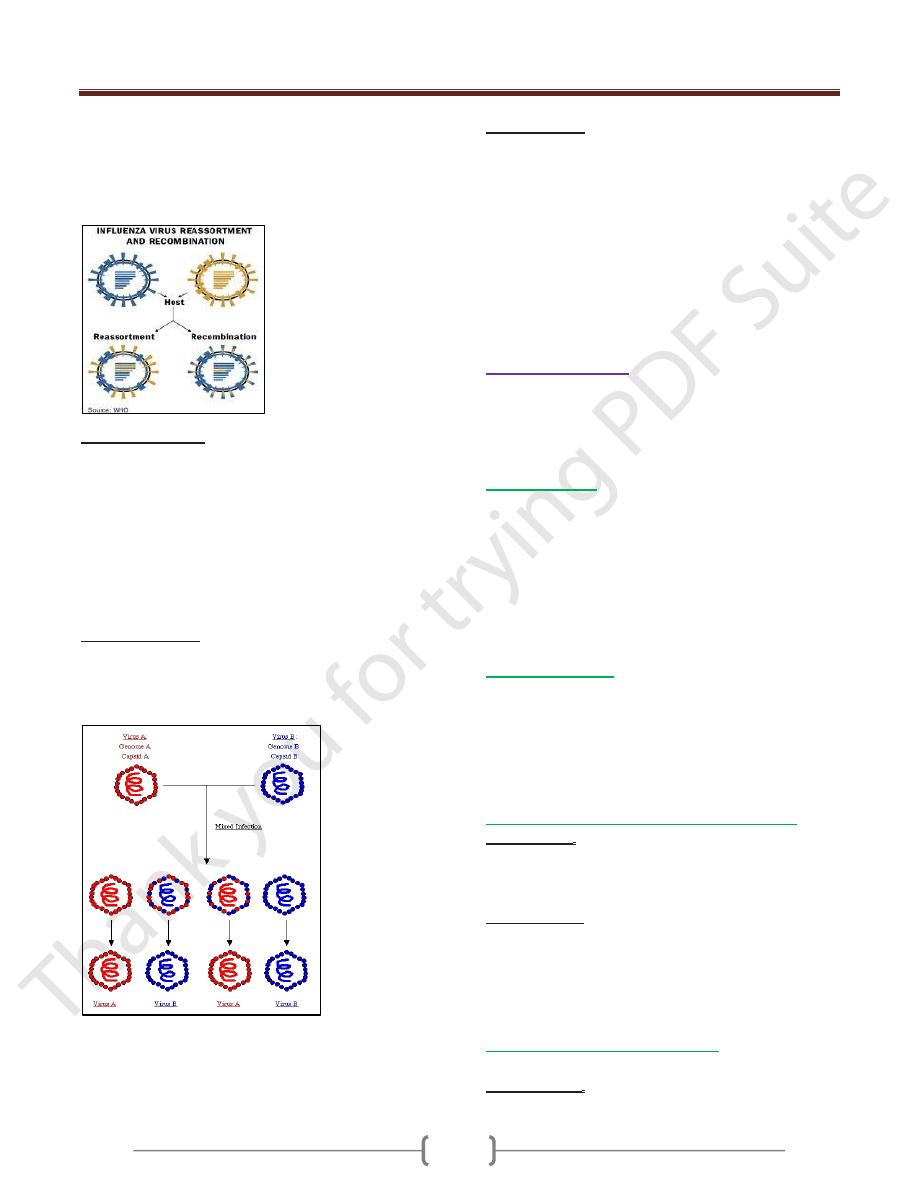
Unit 4: Virology
255
base sequence homology. Recombination can readily be
demonstrated for viruses with DS DNA as well as by
RNA virus but with a very low frequency.
In case of segmented genome virus,
Reassortment
occurs
when an exchange between segments occur.
2) Complementation :
Refers to interaction of viral gene products in cells
infected with two viruses, one or both of which may be
defective. It results in the replication of one or both under
conditions in which replication would not ordinarily
occur. One of the virus provides a gene product in which
the second is defective, allowing the second virus to grow.
The genotype of the two viruses remain unchanged. if
both mutants are defective in the same gene product, they
will not be able to complement each others growth.
3) Phenotypic mixing
The genome of virus type A can be coated with the
surface proteins of virus type B. This phenotypically virus
can infect cells as determined by its type B protein coat
but the progeny virus from this infection has a type A coat
Pseudotypes, which consist of the nucleocapsid of one
virus and the envelope of another e.g nucleocapsid of
vesicular stomatitis virus and the envelope of HIV are
currently being used in study immune response to HIV.
D- Interference
Infection of either cell cultures or whole animals with two
viruses often leads to an inhibition of multiplication of
one of the viruses.
Several mechanisms as a cause of interference:
1) One virus may inhibit the ability of the second to adsorb
to the cell.
2) One virus may compete for the second for the component
of the replication apparatus.
3) The first virus may cause the infected cell to produce an
inhibitor that prevents replication of the second virus.
Viral pathogenesis
The ability of viruses to cause disease can be viewed on
two distinct levels: (1) the changes that occur within
individual cells (2) the process that takes place in the
infected patient.
The infected cells
Four main effects of virus infection on the cell
1) death
2) Fusion of the cells to form multinucleated giant cells
or inclusion bodies (discrete areas containing viral
proteins or viral particles and may be intracytoplasmic
or intranuclear).
3) Malignant transformation.
4) No apparent morphologic or functional changes.
The infected patient
Pathogenesis in the infected patient involves:
1) Transmission of virus and its entry into the host.
2) Replication of the virus.
3) Spread of the virus to other cells and organs.
4) Immune response.
5) Persistence of the virus.
Transmission of virus and its entry into the host.
Transmission. Viruses can be transmitted horizontally
(within a group of individuals or vertically (from mother
to offspring). Vertical infection is either transovarial or by
infection of the virus in utero (ascending or diaplacental).
Portal of entry. The most important portals of entry for
viruses are themucosa of the respiratory and
gastrointestinal tracts. Intact epidermispresents a barrier to
viruses, which can, however, be overcome through
microtraumata (nearly always present) or mechanical
inoculation (e.g., bloodsucking arthropods).
Viral dissemination in the organism.
There are two forms of infection:
Local infection. In this form of infection, the viruses
spread only from cell to cell. The infection and manifest

Unit 4: Virology
251
disease are thus restricted to the tissues in the immediate
vicinity of the portal of entry. Example: rhinoviruses that
reproduce only in the cells of the upper respiratory tract.
Generalized infection. In this type, the viruses usually
replicate to some extent at the portal of entry and are then
disseminated via the lymph ducts or bloodstream and
reach their target organ either directly or after infecting a
further organ. When the target organ is reached, viral
replication and the resulting cell destruction become so
widespread that clinical symptoms develop. Examples of
such infection courses are seen with enteroviruses that
replicate mainly in the intestinal epithelium, but cause no
symptoms there.
Clinical symptoms in these infections first arise in the
target organs such as the CNS (polioviruses, echoviruses)
or musculature (coxsackie viruses).
Another mode of viral dissemination in the
macroorganism is neurogenic spread along the nerve
tracts, from the portal of entry to the CNS (rabies), or in
the opposite direction from the ganglions where the
viruses persist in a latent state to the target organ (herpes
simplex).
Virus excretion
Excretion of newly produced viruses depends on the
localization of viral replication. For example, viruses that
infect the respiratory tract are excreted in expired air
(droplet infection).
In generalized infections not only the target organ is
involved in excretion, but that primary viral replication at
the portal of entry also contributes to virus excretion (for
example enteroviruses, which replicate primarily in the
intestinal wall and are excreted in feces).
It is important to know, patients are contagious before
they really become ill because excretion of new virus
progeny precede the onset of illness.
Persistent viral infections
In most viral infections, the virus doesn’t remain in the
body for a significant period after clinical recovery, but in
certain instances, the virus persists for long periods either
intact or in the form of subviral component (e.g.: the
genome).
There are three types of persistent viral infections:
1) Chronic carrier infection
: refers to people who produce
virus for long periods of time and can serve as a source of
infection for others.
2) Latent infections
: are those infections that are not
producing virus at the present time but can be reactivated
at subsequent time e.g latent infections that are associated
with herpes simplex virus infection.
3) Slow virus infections
: refer to those diseases with a long
incubation period often in years. Some are caused by
virus (progressive multifocal leukoencephalopathy) ,
whereas others are caused by prions ( Creutzfeldt-Jakob
disease).
Evasion of host defenses:
Viruses have several ways to evade host defense:
1) Some viruses encode the receptors for various mediators
of immunity e,g: Vaccina virus encode protein that bind
to IL-1. Fibroma virus encodes protein that binds to TNF.
These virus encoded proteins called Cytokine decoy.
2) Human Iimmunodeficiency V, Herpes v, CMV reduce
Class I MHC expression.
3) HIV,Epstein Barre V,Adeno v. synthesis RNAs that block
phosphrylation of an initiation factor (eIF2) which reduce
ability of INF to block viral replication
4) CMV encodes a micro RNA binds to mRNA of a cell
surface ligand for NK cell.
5) Measles block IL-12.
6) virus has multiple antigenic type
e.g Rhinovirus (100 serotype).