the muscles of the hands and the muscles of speech. Point stimulation in these
human beings who were undergoing neurosurgical operations. Note that more
the different muscle areas as mapped by Penfield and Rasmussen. This mapping
more graphically in Figure 55–2, which shows the degrees of representation of
the longitudinal fissure. This topographical organization is demonstrated even
and the leg and foot areas, in the part of the primary motor cortex that dips into
midportion of the primary motor cortex; the trunk, near the apex of the brain;
face and mouth region near the sylvian fissure; the arm and hand area, in the
ferent muscle areas of the body in the primary motor cortex, beginning with the
Figure 55–1 lists the approximate topographical representations of the dif-
classification of the brain cortical areas, shown in Figure 47–5.)
deep into the longitudinal fissure. (This area is the same as area 4 in Brodmann’s
fissure, spreads superiorly to the uppermost portion of the brain, and then dips
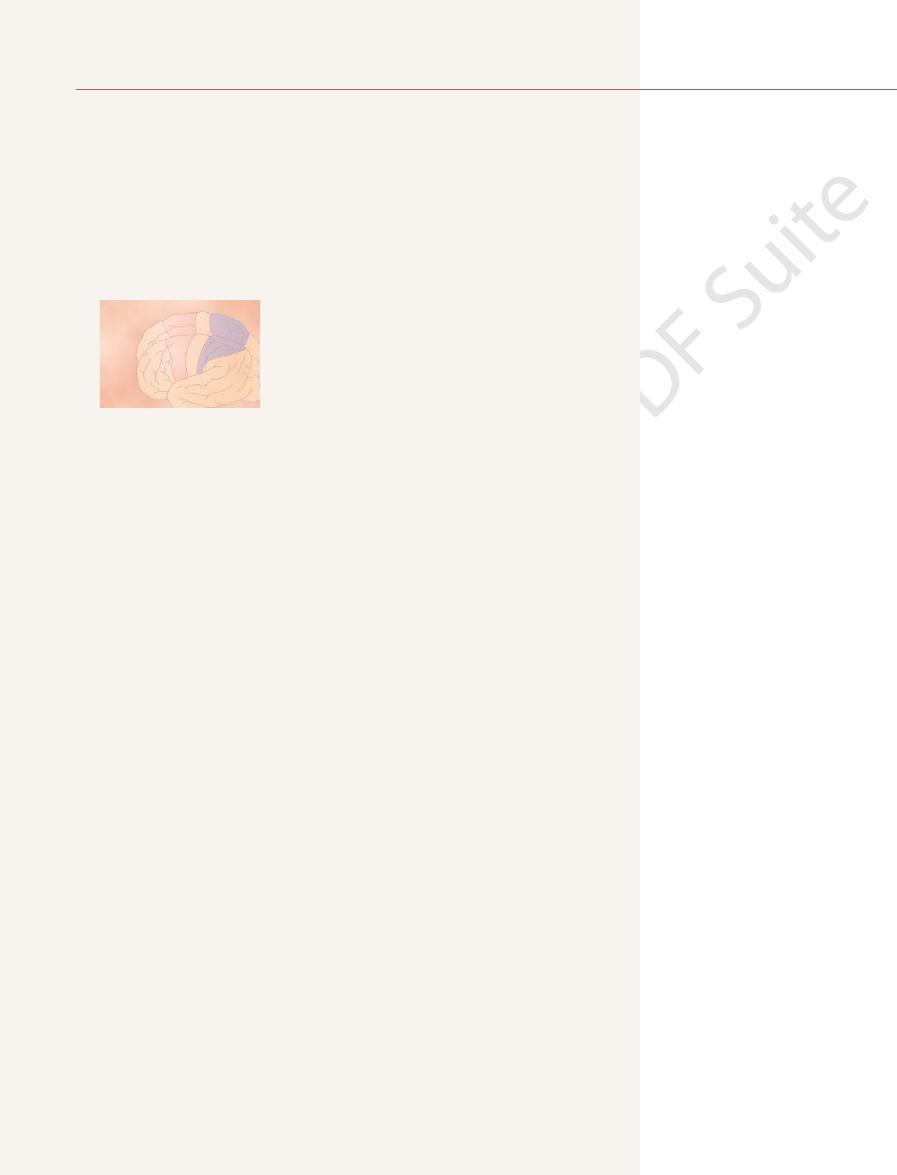
the frontal lobes anterior to the central sulcus. It begins laterally in the sylvian
The primary motor cortex, shown in Figure 55–1, lies in the first convolution of
tions: (1) the
The motor cortex itself is divided into three subareas, each of which has its
feeds the motor cortex many of the signals that initiate motor activities.
(an area discussed in detail in earlier chapters), which
Posterior to the central sulcus is the
the frontal lobes, is the
central cortical sulcus, occupying approximately the posterior one third of
Figure 55–1 shows the functional areas of the cerebral cortex. Anterior to the
MOTOR CORTEX AND CORTICOSPINAL TRACT
ments of the fingers and hands. This chapter and Chapter 56 explain the inter-
centers on the way. This is especially true for control of the fine dexterous move-
pathway to the anterior motor neurons of the cord, bypassing some motor
For a few types of movements, however, the cortex has almost a direct
control signals to the muscles.
bellum. These lower centers, in turn, send specific
areas—the cord, brain stem, basal ganglia, and cere-
vates “patterns” of function stored in lower brain
Most “voluntary” movements initiated by the
In this chapter, we discuss control of body move-
C
H
A
P
T
E
R
5
5
685
Cortical and Brain Stem Control
of Motor Function
ments by the cerebral cortex and brain stem.
cerebral cortex are achieved when the cortex acti-
play among the different motor areas of the brain and spinal cord to provide
overall synthesis of voluntary motor function.
motor cortex.
somatosensory cortex
own topographical representation of muscle groups and specific motor func-
primary motor cortex, (2) the premotor area, and (3) the supple-
mentary motor area.
Primary Motor Cortex
was done by electrically stimulating the different areas of the motor cortex in
than one half of the entire primary motor cortex is concerned with controlling
hand and speech motor areas on rare occasion causes contraction of a single
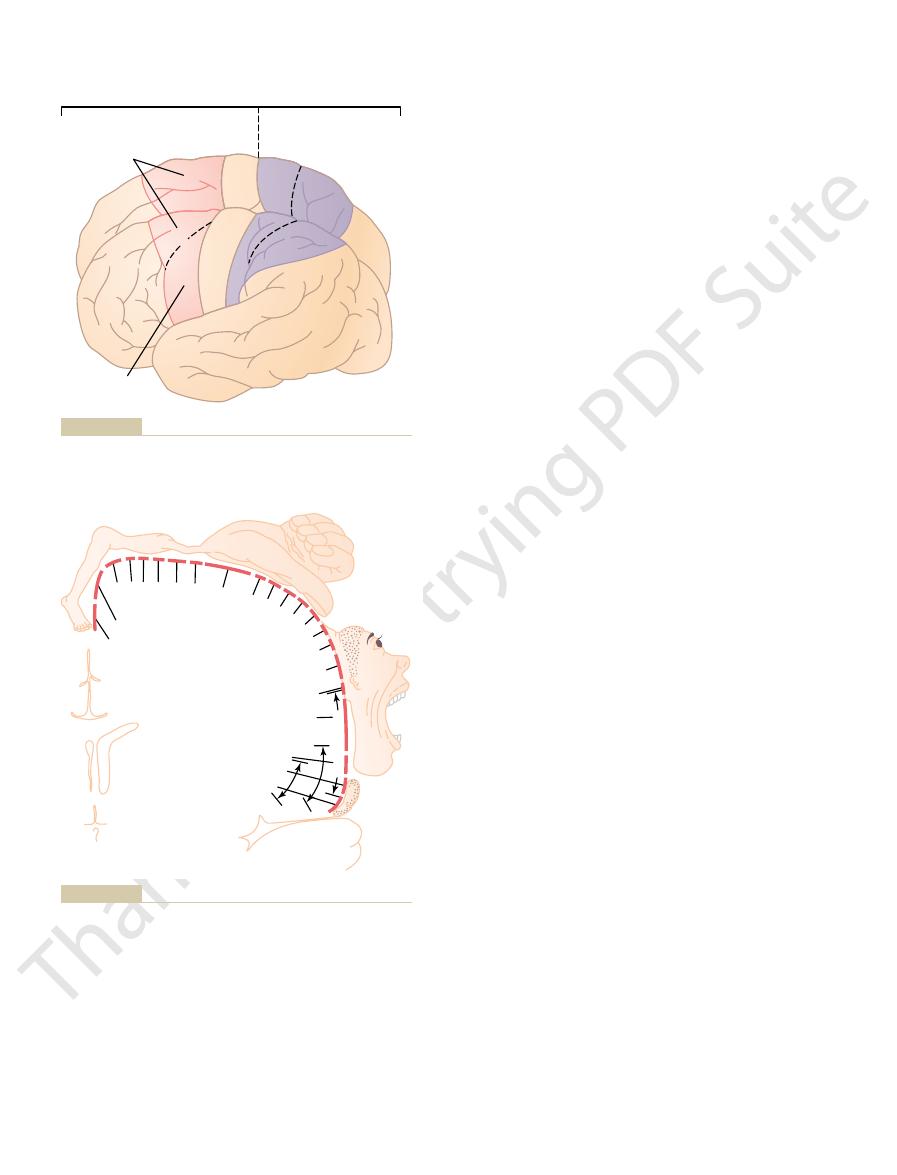
Figure 55–3) that control specific motor functions.
Motor Cortex
Control Found in the Human
Some Specialized Areas of Motor
movements of the head and eyes, and so forth, as back-
of the different segments of the body, positional
body-wide attitudinal movements, fixation movements
tions required for climbing. In general, this area func-
movements of both hands simultaneously; these
are often bilateral rather than unilateral. For instance,
cortex. Contractions elicited by stimulating this area
tion. It lies mainly in the longitudinal fissure but
The supplementary motor area has yet another topo-
Supplementary Motor Area
coordinated muscle activity.
amus, and primary motor cortex constitute a complex
cortex. Thus, the premotor cortex, basal ganglia, thal-
to excite specific muscles or, often, by way of the basal
image.This posterior part of the premotor cortex sends
rior premotor cortex, this image excites each succes-
movement that is to be performed. Then, in the poste-
first develops a “motor image” of the total muscle
results, the most anterior part of the premotor area
oriented to perform specific tasks. To achieve these
cortex. For instance, the pattern may be to position
much more complex “patterns” of movement than
moves upward, the hand, arm, trunk, and leg areas are
mouth and face areas located most laterally; as one
same as that of the primary motor cortex, with the
similar to those of the premotor area. The topograph-
the supplementary motor area, which has functions
riorly into the longitudinal fissure, where it abuts
The premotor area, also shown in Figure 55–1, lies 1
To do this, it excites a “pattern” of separate muscles,
a specific movement rather than one specific muscle.
muscles instead. To express this in another way, exci-
muscle; most often, stimulation contracts a group of
The Nervous System: C. Motor and Integrative Neurophysiology
686
Unit XI
tation of a single motor cortex neuron usually excites
each of which contributes its own direction and
strength of muscle movement.
Premotor Area
to 3 centimeters anterior to the primary motor cortex,
extending inferiorly into the sylvian fissure and supe-
ical organization of the premotor cortex is roughly the
encountered.
Nerve signals generated in the premotor area cause
the discrete patterns generated in the primary motor
the shoulders and arms so that the hands are properly
sive pattern of muscle activity required to achieve the
its signals either directly to the primary motor cortex
ganglia and thalamus back to the primary motor
overall system for the control of complex patterns of
graphical organization for the control of motor func-
extends a few centimeters onto the superior frontal
stimulation frequently leads to bilateral grasping
movements are perhaps rudiments of the hand func-
tions in concert with the premotor area to provide
ground for the finer motor control of the arms and
hands by the premotor area and primary motor cortex.
Neurosurgeons have found a few highly specialized
motor regions of the human cerebral cortex (shown in
Motor
Sensory
Primary
motor
cortex
Supplementary
area
Premotor
area
Face
Hand
Arm
4
Trunk
Feet
Legs
7
6
5
3, 2, 1
Mouth
Somatic
area 1
Somatic
association
area
The numbers 4, 5, 6, and 7 are Brodmann’s cortical areas, as
Motor and somatosensory functional areas of the cerebral cortex.
Figure 55–1
explained in Chapter 47.
Trunk
Shoulder
Elbow
Wrist
Hand
Little finger
Ring finger
Middle
finger
Index finger
Thumb
Neck
Brow
Face
Lips
Vocali
zation
Mastication
Salivation
Jaw
Tongue
Swallow
ing
Eyel
id and eyeball
Hip
Knee
Ankle
Toes
tion. New York: Hafner, 1968.)
Cerebral Cortex of Man: A Clinical Study of Localization of Func-
the motor cortex. (Redrawn from Penfield W, Rasmussen T: The
Degree of representation of the different muscles of the body in
Figure 55–2

corticospinal tract is more than 1 million, so these large
corticospinal tract. The total number of fibers in each
are about 34,000 of these large Betz cell fibers in each
sion of any signals from the brain to the cord. There
ity of about 70 m/sec, the most rapid rate of transmis-
are about 60 micrometers in diameter, and their fibers
found only in the primary motor cortex. The Betz cells
Betz cells,
giant pyramidal cells,
diameter of 16 micrometers. These fibers originate
The most impressive fibers in the pyramidal tract are
These fibers may be concerned with control of bilat-
ventral corticospinal tracts.
in the dorsal horn, and a very few terminate directly
gray matter; a few terminate on sensory relay neurons
of the cord, finally terminating principally on the
The majority of the
pyramids of the medulla.
and then downward through the brain stem, forming
After leaving the cortex, it passes through the pos-
somatosensory areas posterior to the central sulcus.
mentary motor areas, and 40 per cent from the
cortex, 30 per cent from the premotor and supple-
shown in Figure 55–4. The corticospinal tract
pyram-
The most important output pathway from the motor
Corticospinal (Pyramidal) Tract
particularly the hands and fingers.
ments, especially of the distal segments of the limbs,
In general, the direct pathways
basal ganglia, cerebellum,
Motor Cortex to the Muscles
Transmission of Signals from the
ful, a condition called
other lesions cause destruction in this area, hand
important for “hand skills.” That is, when tumors or
field; it directs the head toward different objects.
This area is closely associated with the eye movement
ation area, electrical stimulation elicits head rotation.
movements such as blinking.
in Chapter 51. This frontal area also controls eyelid
by signals from the occipital visual cortex, as explained
involuntarily onto specific objects, an effect controlled
toward different objects. Instead, the eyes tend to lock
ling voluntary eye movements. Damage to this area
immediately above Broca’s area is a locus for control-
“Voluntary” Eye Movement Field.
during speech. Thus, the premotor neuronal activities
causes appropriate respiratory function, so that respi-
“no” or “yes.” A closely associated cortical area also
vocalizing, but it does make it impossible for the
above the sylvian fissure. This region is called
area labeled “word formation” lying immediately
Figure 55–3 shows a premotor
Broca’s Area and Speech.
following.
areas. Some of the more important regions are the
These regions have been localized either by electrical
Chapter 55
Cortical and Brain Stem Control of Motor Function
687
stimulation or by noting the loss of motor function
when destructive lesions occur in specific cortical
anterior to the primary motor cortex and immediately
Broca’s
area. Damage to it does not prevent a person from
person to speak whole words rather than uncoordi-
nated utterances or an occasional simple word such as
ratory activation of the vocal cords can occur simulta-
neously with the movements of the mouth and tongue
related to speech are highly complex.
In the premotor area
prevents a person from voluntarily moving the eyes
Head Rotation Area.
Slightly higher in the motor associ-
Area for Hand Skills.
In the premotor area immediately
anterior to the primary motor cortex for the hands and
fingers is a region neurosurgeons have identified as
movements become uncoordinated and nonpurpose-
motor apraxia.
Motor signals are transmitted directly from the cortex
to the spinal cord through the corticospinal tract and
indirectly through multiple accessory pathways that
involve the
and various
nuclei of the brain stem.
are concerned more with discrete and detailed move-
cortex is the corticospinal tract, also called the
idal tract,
originates about 30 per cent from the primary motor
terior limb of the internal capsule (between the
caudate nucleus and the putamen of the basal ganglia)
the
pyramidal fibers then cross in the lower medulla to the
opposite side and descend into the lateral corticospinal
tracts
interneurons in the intermediate regions of the cord
on the anterior motor neurons that cause muscle
contraction.
A few of the fibers do not cross to the opposite side
in the medulla but pass ipsilaterally down the cord in
the
Many if not most of
these fibers eventually cross to the opposite side of the
cord either in the neck or in the upper thoracic region.
eral postural movements by the supplementary motor
cortex.
a population of large myelinated fibers with a mean
from
called
that are
transmit nerve impulses to the spinal cord at a veloc-
Chewing
Swallowing
Tongue
Jaw
Vocalization
Lips
Neck
Finge
rs
Hips
Legs
Feet
Thumb
Trunk
Arms
Supplemental
and premotor
areas
Primary
motor
cortex
Contralateral
eye movements
Head rotation
Choice
of words
Eye
fixation
Hand skills
Word formation
(Broca’s area)
cortex and location of other cortical areas responsible for specific
Representation of the different muscles of the body in the motor
Figure 55–3
types of motor movements.

located in the mesencephalon, func-
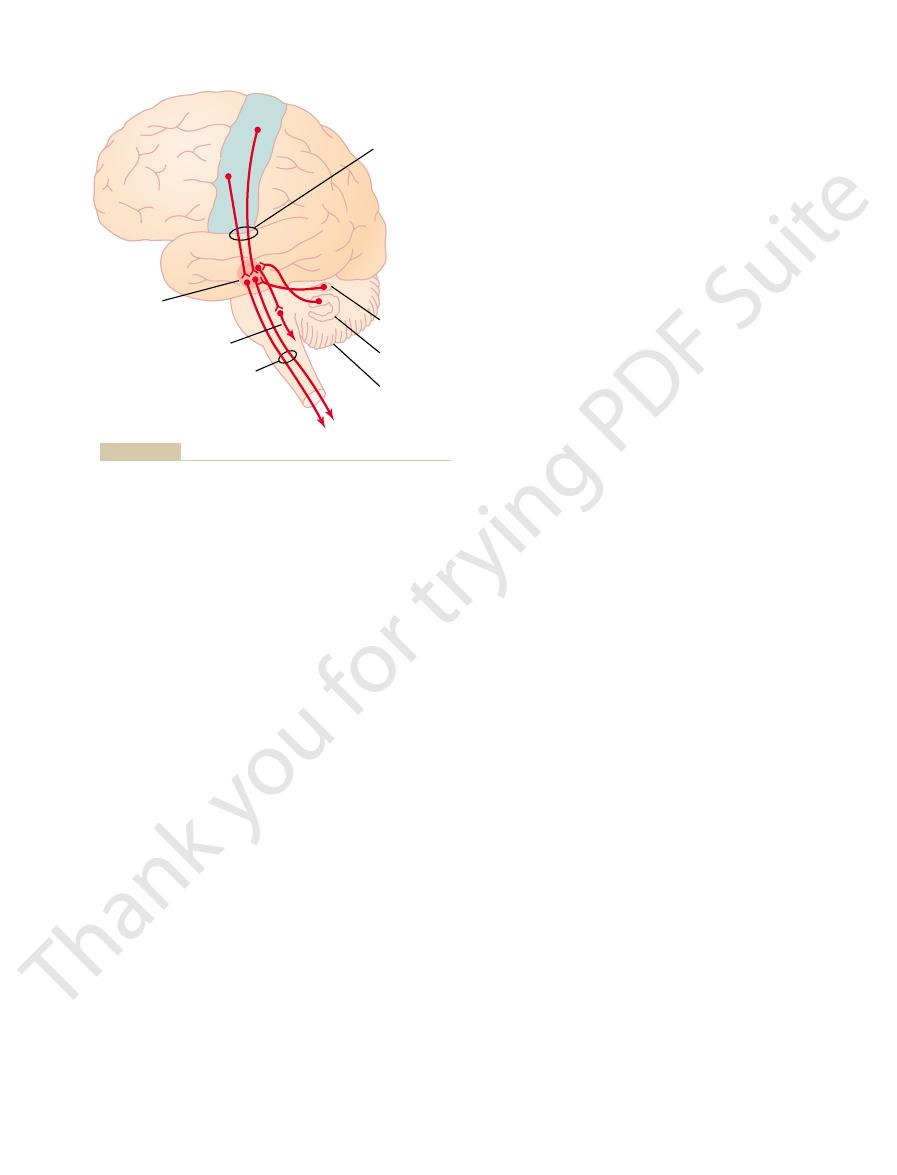
red nucleus,
The
Signals to the Spinal Cord
Pathway for Transmitting Cortical
Red Nucleus Serves as an Alternative
These fibers control the general level of excitability
5. Fibers from the intralaminar nuclei of the thalamus.
cortex, basal ganglia, and cerebellum.
from the cerebellum and basal ganglia. These tracts
nuclei of the thalamus, which in turn receive signals
4. Tracts from the ventrolateral and ventroanterior
muscle signals from the peripheral body.
ventrobasal complex of the thalamus. These relay
3. Somatosensory fibers that arrive directly from the
These fibers connect corresponding areas of the
callosum from the opposite cerebral hemisphere.
2. Subcortical fibers that arrive through the corpus
cortices.
the motor cortex, and (c) the visual and auditory
somatosensory areas of the parietal cortex, (b) the
the cerebral cortex, especially from (a) the
1. Subcortical fibers from adjacent regions of
ate course of motor action. The more important incom-
received, the motor cortex operates in association with
as hearing and vision. Once the sensory information is
also, to some degree, from other sensory systems such
The functions of the motor cortex are controlled mainly
Incoming Fiber Pathways to the
spinal cord to cause a motor activity.
Thus, the basal ganglia, brain stem, and cerebellum all
and from there, secondary
6. Collaterals also terminate in the
cerebellar hemispheres.
pontocerebellar fibers,
in the pontile nuclei, which give rise to the
5. A tremendous number of motor fibers synapse
vestibulocerebellar tracts.
vestibulospinal tracts,
brain stem; from there, signals go to the cord by
4. A moderate number of motor fibers deviate into
From these, additional fibers
3. A moderate number of motor fibers pass to
contractions.
chapter, mainly to control body postural muscle
stem and spinal cord, as discussed in the next
there, additional pathways extend into the brain
From
2. A large number of fibers pass from the motor
“sharpening” the boundaries of the excitatory
of the cortex when the Betz cells discharge, thereby
collaterals back to the cortex itself. These
1. The axons from the giant Betz cells send short
cerebrum and brain stem, including the following:
tional, mainly small fibers that go to deep regions in the
The motor cortex gives rise to large numbers of addi-
fibers represent only 3 per cent of the total. The other
The Nervous System: C. Motor and Integrative Neurophysiology
688
Unit XI
97 per cent are mainly fibers smaller than 4 microme-
ters in diameter that conduct background tonic signals
to the motor areas of the cord.
Other Fiber Pathways from the Motor Cortex
collaterals are believed to inhibit adjacent regions
signal.
cortex into the caudate nucleus and putamen.
red
nuclei of the midbrain.
pass down the cord through the rubrospinal tract.
the reticular substance and vestibular nuclei of the
way of reticulospinal and
and others go to the cerebellum by way of
reticulocerebellar and
carrying signals into the
inferior olivary
nuclei,
olivocerebellar
fibers transmit signals to multiple areas of the
cerebellum.
receive strong motor signals from the corticospinal
system every time a signal is transmitted down the
Motor Cortex
by nerve signals from the somatosensory system but
the basal ganglia and cerebellum to excite an appropri-
ing fiber pathways to the motor cortex are the following:
adjacent areas of the frontal cortex anterior to
cortices in the two sides of the brain.
mainly cutaneous tactile signals and joint and
provide signals that are necessary for coordination
among the motor control functions of the motor
of the motor cortex in the same way they control
the general level of excitability of most other
regions of the cerebral cortex.
tions in close association with the corticospinal tract.
Posterior limb of internal
capsule
Genu of corpus callosum
Longitudinal fascicles
of pons
Motor cortex
Basis pedunculi of
mesencephalon
Pyramid of medulla
oblongata
Lateral corticospinal tract
Ventral corticospinal tract
Pyramidal tract. (Modified from Ranson SW, Clark SL: Anatomy of
Figure 55–4
the Nervous System. Philadelphia: WB Saunders, 1959.)
system, using information from multiple input sources
The neurons of
Function of Each Column of Neurons.
bral cortex itself.
through 4. And the sixth layer gives rise mainly to
versely, the input signals all enter by way of layers 2
in the fifth layer of cells from the cortical surface. Con-
throughout nearly all the cerebral cortex. The pyram-
column has six distinct layers of cells, as is true
times stimulating just a single muscle. Also, each
stimulating a group of synergistic muscles, but some-
Each column of cells functions as a unit, usually
columns a fraction of a millimeter in diameter, with
. In like manner,
In Chapters 47 and 51, we pointed out that the
Vertical Columnar Arrangement of the Neurons in the Motor
Control Areas by the Primary Motor
Excitation of the Spinal Cord Motor
less often both clinically and physiologically.
For this reason, the term “extrapyramidal” is being used
tions to the so-called extrapyramidal system as a whole.
nuclei, and often the red nuclei. This is such an all-
reticular formation of the brain stem, the vestibular
These include pathways through the basal ganglia, the
extrapyramidal motor system
The term
medial motor system of the cord,
lospinal system, which lies mainly medially in the cord
of the cord,
the limbs. Therefore, the corticospinal and rubrospinal
cospinal tract, and terminates on the interneurons and
columns of the spinal cord, along with the corti-
system. Further, the rubrospinal tract lies in the lateral
Therefore, the pathway through the red nucleus to
impaired. Wrist movements are still functional, which
can still occur, except that the movements for fine
corubrospinal pathway is intact, discrete movements
signals from the motor cortex to the spinal cord. When
The corticorubrospinal pathway serves as an acces-
motor cortex. This is especially true in human beings,
muscles. However, the fineness of representation of
of the motor cortex. Therefore, stimulation of a single
representation of all the muscles of the body, as is true
The magnocel-
cerebellum, similar to the connections between the
The red nucleus also has close connections with the
motor neurons, along with some corticospinal fibers.
matter, along with the corticospinal fibers, but some of
The rubrospinal fibers terminate mostly on the
the motor cortex. These large neurons then give rise to
the red nucleus, the
cephalon. These fibers synapse in the lower portion of
As shown in Figure 55–5, it receives a large number of
Chapter 55
Cortical and Brain Stem Control of Motor Function
689
direct fibers from the primary motor cortex through
the corticorubral tract, as well as branching fibers from
the corticospinal tract as it passes through the mesen-
magnocellular portion, which con-
tains large neurons similar in size to the Betz cells in
the rubrospinal tract, which crosses to the opposite
side in the lower brain stem and follows a course
immediately adjacent and anterior to the corticospinal
tract into the lateral columns of the spinal cord.
interneurons of the intermediate areas of the cord gray
the rubrospinal fibers terminate directly on anterior
motor cortex and the cerebellum.
Function of the Corticorubrospinal System.
lular portion of the red nucleus has a somatographic
point in this portion of the red nucleus causes con-
traction of either a single muscle or a small group of
the different muscles is far less developed than in the
who have relatively small red nuclei.
sory route for transmission of relatively discrete
the corticospinal fibers are destroyed but the corti-
control of the fingers and hands are considerably
is not the case when the corticorubrospinal pathway is
also blocked.
the spinal cord is associated with the corticospinal
motor neurons that control the more distal muscles of
tracts together are called the lateral motor system
in contradistinction to a vestibuloreticu-
and is called the
as
discussed later in this chapter.
“Extrapyramidal” System
is widely used in
clinical circles to denote all those portions of the brain
and brain stem that contribute to motor control but are
not part of the direct corticospinal-pyramidal system.
inclusive and diverse group of motor control areas that
it is difficult to ascribe specific neurophysiologic func-
Cortex and Red Nucleus
Cortex.
cells in the somatosensory cortex and visual cortex are
organized in vertical columns of cells
the cells of the motor cortex are organized in vertical
thousands of neurons in each column.
idal cells that give rise to the corticospinal fibers all lie
fibers that communicate with other regions of the cere-
each column operate as an integrative processing
Motor
cortex
Interpositus
nucleus
Dentate
nucleus
Cerebellum
Red nucleus
Reticular formation
Rubrospinal tract
Corticorubral
tract
relation of this pathway to the cerebellum.
Corticorubrospinal pathway for motor control, showing also the
Figure 55–5

direct stimulation by the corticospinal fibers.
assist” stimulation of the muscle, in addition to the
skeletal muscle fibers do, thus eliciting reflex “servo-
movements initiated from the brain, and probably also
all times, helping to damp any oscillations of the motor
brain. For example, the stretch reflex is functional at
same patterns are also important when the cord’s ante-
response to sensory nerve stimulation. Many of these
Chapter 54, recall that the spinal cord can provide
From
finger, and thumb actions.
muscle contraction. This is in keeping with the fact that
nate directly on the anterior motor neurons, thus
hands and fingers are represented, large numbers of
area of the cord gray matter.
portions of the lateral white columns. Their fibers ter-
middle of the anterior horn gray matter. The corti-
Figure 55–6 shows a cross section of a spinal cord
Stimulation of the Spinal Motor Neurons
tightness of the hand grasp.
excitation of the muscles and, therefore, increase the
the skin receptors can, if necessary, cause further
around an object being grasped, the signals from
against an object, such as compression of the fingers
spindles. In the case of the tactile receptors, if the
idal cells further excite the muscle, helping its con-
muscle fibers have not contracted enough. The pyram-
become stretched and, therefore, excited. Signals from
fibers contract, the central portions of the spindles
muscle spindles, if the fusimotor muscle fibers in the
contraction in the following ways: In the case of the
overlying the muscles. These somatic signals often
muscle tendons, or (3) the tactile receptors of the skin
the muscle spindles, (2) the tendon organs of the
action. Most of these somatosensory signals arise in (1)
muscle to contract, somatosensory signals return all
When nerve signals from the motor cortex cause a
Cortex Helps Control the Precision of
Somatosensory Feedback to the Motor
the next chapter.
rapid initiation of muscle contraction, as explained in
bellum, and the cerebellum plays an important role in
in the primary motor cortex. This may be related to the
dynamic and static characteristics, except that a
The neurons of the red nucleus have similar
rate, but they continue firing at this slow rate to
Then the static neurons fire at a much slower
of force.
of a contraction, causing the initial rapid
The dynamic neurons are
static neurons.
of pyramidal cell neurons, one called
do this, each column of cells excites two populations
excitation is provided to cause muscle contractions. To
periods thereafter. This is the usual manner in which
to cause initial rapid contraction, then a much weaker
Dynamic and Static Signals Transmitted by the Py-
Usually, 50 to 100 pyramidal cells need to be excited
single pyramidal cell can seldom excite a muscle.
neously. This is important, because stimulation of a
addition, each column can function as an amplifying
to determine the output response from the column. In
The Nervous System: C. Motor and Integrative Neurophysiology
690
Unit XI
system to stimulate large numbers of pyramidal fibers
to the same muscle or to synergistic muscles simulta-
simultaneously or in rapid succession to achieve defin-
itive muscle contraction.
ramidal Neurons.
If a strong signal is sent to a muscle
continuing signal can maintain the contraction for long
dynamic neurons
and the other
excessively excited for a short period at the beginning
development
main-
tain the force of contraction as long as the contraction
is required.
greater percentage of dynamic neurons is in the red
nucleus and a greater percentage of static neurons is
fact that the red nucleus is closely allied with the cere-
Muscle Contraction
the way from the activated region of the body to the
neurons in the motor cortex that are initiating the
cause positive feedback enhancement of the muscle
spindles contract more than the large skeletal muscle
these spindles then return rapidly to the pyramidal
cells in the motor cortex to signal them that the large
traction to catch up with the contraction of the muscle
muscle contraction causes compression of the skin
segment demonstrating (1) multiple motor and senso-
rimotor control tracts entering the cord segment and
(2) a representative anterior motor neuron in the
cospinal tract and the rubrospinal tract lie in the dorsal
minate mainly on interneurons in the intermediate
In the cervical enlargement of the cord where the
both corticospinal and rubrospinal fibers also termi-
allowing a direct route from the brain to activate
the primary motor cortex has an extremely high
degree of representation for fine control of hand,
Patterns of Movement Elicited by Spinal Cord Centers.
certain specific reflex patterns of movement in
rior motor neurons are excited by signals from the
providing at least part of the motive power required
to cause muscle contractions when the intrafusal fibers
of the muscle spindles contract more than the large
Sensory neurons
Propriospinal tract
Interneurons
Corticospinal tract
from pyramidal cells
of cortex
Rubrospinal tract
Reticulospinal tract
Anterior motor
neuron
Motor nerve
Tectospinal and
reticulospinal tracts
Vestibulospinal and
reticulospinal tracts
motor neurons.
Convergence of different motor control pathways on the anterior
Figure 55–6

Figure 55–7 shows the locations of the reticular and
Vestibular Nuclei
Roles of the Reticular and
Support of the Body Against Gravity
stem’s
following sections, we discuss the role of the brain stem
“command signals” from higher neural centers. In the
Finally, the brain stem serves as a way station for
6. Control of eye movements
5. Control of equilibrium
4. Control of many stereotyped movements of the
3. Partial control of gastrointestinal function
2. Control of the cardiovascular system
1. Control of respiration
special control functions, such as the following:
tions from the neck down. But in another sense, the
spinal cord upward into the cranial cavity, because it
In one sense, it is an extension of the
medulla, pons,
The brain stem consists of the
Controlling Motor Function
human being.
spasticity that normally accompanies a “stroke” in a
we discuss more fully later in the chapter. This is the
cause excessive spastic tone in the involved muscles, as
inhibited”), they become spontaneously active and
these nuclei cease their state of inhibition (i.e., are “dis-
vestibular and reticular brain stem motor nuclei. When
motor cortex. These pathways normally inhibit the
side). This spasm results mainly from damage to acces-
instances,
cent parts of the brain such as the basal ganglia. In these
of the motor cortex, especially those caused by a
results. Most lesions
the motor neurons of the spinal cord; when this stimu-
The primary motor cortex
and fingers.
of finely controlled movements, especially of the hands
From these observations, one can conclude that
gone.
ability to control the fine movements is
tract; rather, the
This does not mean
especially of the hands and fingers.
discrete movements of the distal segments of the limbs,
can still occur, but there is
damaged, gross postural and limb “fixation” movements
muscles. If the sublying caudate nucleus and adjacent
Removal of the Primary Motor Cortex (Area Pyramidalis).
the caudate nucleus and the putamen.Also, experiments
supplying the brain. In either case, the result is loss of
common abnormality called a “stroke.” This is caused
The motor control system can be damaged by the
the body.
normal motor activities, particularly for such functions
“command” signals from the brain. Thus, simple
postural mechanisms, can each be activated by
withdrawal, stepping and walking, scratching, and
Finally, other cord reflex mechanisms, such as
antagonistic pairs of muscles.
the antagonist muscle at the same time; this is achieved
Also, when a brain signal excites a muscle, it usually
Chapter 55
Cortical and Brain Stem Control of Motor Function
691
is not necessary to transmit an inverse signal to relax
by the reciprocal innervation circuit that is always
present in the cord for coordinating the function of
command signals from the brain can initiate many
as walking and attaining different postural attitudes of
Effect of Lesions in the Motor Cortex or in the Corticospinal
Pathway—The “Stroke”
either by a ruptured blood vessel that hemorrhages into
the brain or by thrombosis of one of the major arteries
blood supply to the cortex or to the corticospinal tract
where it passes through the internal capsule between
have been performed in animals to selectively remove
different parts of the motor cortex.
Removal of a portion of the primary motor cortex—the
area that contains the giant Betz pyramidal cells—
causes varying degrees of paralysis of the represented
premotor and supplementary motor areas are not
loss of voluntary control of
that the hand and finger muscles themselves cannot con-
the area pyramidalis is essential for voluntary initiation
Muscle Spasticity Caused by Lesions That Damage Large Areas
Adjacent to the Motor Cortex.
normally exerts a continual tonic stimulatory effect on
latory effect is removed, hypotonia
stroke,
involve not only the primary motor cortex but also adja-
muscle spasm almost invariably occurs in the
afflicted muscle areas on the opposite side of the body
(because the motor pathways cross to the opposite
sory pathways from the nonpyramidal portions of the
Role of the Brain Stem in
and mes-
encephalon.
contains motor and sensory nuclei that perform motor
and sensory functions for the face and head regions in
the same way that the spinal cord performs these func-
brain stem is its own master because it provides many
body
in controlling whole-body movement and equilibrium.
Especially important for these purposes are the brain
reticular nuclei and vestibular nuclei.
—
vestibular nuclei in the brain stem.
Pontine reticular
nuclei
Medullary reticular
nuclei
Vestibular nuclei
Locations of the reticular and vestibular nuclei in the brain stem.
Figure 55–7

The vestibular apparatus, shown in Figure 55–9, is the
Vestibular Apparatus
Vestibular Sensations and
occur in other neuromotor diseases, especially lesions of
develops. We shall see later that other causes of rigidity
ity of the pontine excitatory system occurs, and rigidity
inhibitor system becomes nonfunctional; full overactiv-
ganglia. Lacking this input, the medullary reticular
from the cerebral cortex, the red nuclei, and the basal
The cause of decerebrate rigidity is blockage of nor-
the extensors of the legs.
This rigidity does not occur
decerebrate rigidity.
system are left intact, the animal develops a condition
the midlevel of the mesencephalon, but the pontine and
When the brain stem of an animal is sectioned below
discuss this more fully later in the chapter.
response to signals from the vestibular apparatus.
The specific role of the vestibular nuclei, however,
gravity muscles.
the vestibular nuclei, the pontine reticular system
cord, as shown in Figure 55–8. Without this support of
control the antigravity muscles. The vestibular nuclei
, shown in Figure 55–7, func-
Role of the Vestibular Nuclei to Excite the
special motor activities. The excitatory and inhibitory
At other times, excitation of the medullary reticular
wishes to excite the pontine system to cause standing.
“disinhibit” the medullary system when the brain
Yet some signals from higher areas of the brain can
mally tense.
normal conditions, the body muscles are not abnor-
signals from the pontine reticular system, so that under
These normally activate the medullary reticular
the rubrospinal tract, and (3) other motor pathways.
input collaterals from (1) the corticospinal tract, (2)
55–8. The medullary reticular nuclei receive strong
lateral column of the cord, as also shown in Figure
tract, the
The medullary reticular
Medullary Reticular System.
a standing position, supporting the body against
so much so that four-legged animals can be placed in
excitation of antigravity muscles throughout the body,
by the medullary reticular system, it causes powerful
from deep nuclei of the cerebellum. Therefore, when
excitatory signals from the vestibular nuclei, as well as
natural excitability. In addition, they receive strong
The pontine reticular nuclei have a high degree of
muscles of the limbs.
body, which support the body against gravity—that is,
column of the cord, as shown in Figure 55–8. The fibers
The pontine reticular nuclei
medullary relaxing these same muscles.
function mainly antagonistically to each other, with
medially near the midline. These two sets of nuclei
extend through the entire medulla, lying ventrally and
encephalon, and (2)
The reticular nuclei are divided into two major groups:
Pontine and Medullary Reticular Nuclei
Excitatory-Inhibitory Antagonism Between
The Nervous System: C. Motor and Integrative Neurophysiology
692
Unit XI
(1) pontine reticular nuclei, located slightly posteriorly
and laterally in the pons and extending into the mes-
medullary reticular nuclei, which
the pontine exciting the antigravity muscles and the
Pontine Reticular System.
transmit excitatory signals downward into the cord
through the pontine reticulospinal tract in the anterior
of this pathway terminate on the medial anterior
motor neurons that excite the axial muscles of the
the muscles of the vertebral column and the extensor
the pontine reticular excitatory system is unopposed
gravity without any signals from higher levels of the
brain.
nuclei transmit inhibitory signals to the same anti-
gravity anterior motor neurons by way of a different
medullary reticulospinal tract, located in the
inhibitory system to counterbalance the excitatory
system can inhibit antigravity muscles in certain por-
tions of the body to allow those portions to perform
reticular nuclei constitute a controllable system that is
manipulated by motor signals from the cerebral cortex
and elsewhere to provide necessary background
muscle contractions for standing against gravity and to
inhibit appropriate groups of muscles as needed so
that other functions can be performed.
Antigravity Muscles
All the vestibular nuclei
tion in association with the pontine reticular nuclei to
transmit strong excitatory signals to the antigravity
muscles by way of the lateral and medial vestibu-
lospinal tracts in the anterior columns of the spinal
would lose much of its excitation of the axial anti-
is to selectively control the excitatory signals to the dif-
ferent antigravity muscles to maintain equilibrium in
We
The Decerebrate Animal Develops Spastic Rigidity
medullary reticular systems as well as the vestibular
called
in all muscles of the body but does occur in the anti-
gravity muscles—the muscles of the neck and trunk and
mally strong input to the medullary reticular nuclei
the basal ganglia.
Maintenance of Equilibrium
sensory organ for detecting sensations of equilibrium.
It is encased in a system of bony tubes and chambers
Medullary reticulospinal
tract
Lateral vestibulospinal
tract
Pontine reticulospinal tract
Medial vestibulospinal tract
motor neurons that control the body’s axial musculature.
cord to excite
Vestibulospinal and reticulospinal tracts descending in the spinal
Figure 55–8
(solid lines) or inhibit (dashed lines) the anterior

fluid and tissues. The weight of the statoconia bends
The calcified statoconia have a
vestibular nerve.
The bases and sides of the hair cells synapse with
up into the gelatinous layer.
55–10; these project
one of which is shown in Figure
hair cells,
are embedded. Also in the macula are thou-
upright. Conversely, the
horizontal plane
The
saccule, shown in the top diagram of Figure 55–9, is a
Detecting Orientation of the Head with Respect to Gravity.
“Maculae”—Sensory Organs of the Utricle and Saccule for
utricle,
circular canals,
has little to do with equilibrium. However, the
The cochlea is the
saccule.
chambers, the
cochlearis); three
cochlea
labyrinth. It is composed mainly of the
The top of Figure 55–9 shows the membranous
part of the vestibular apparatus.
The membranous labyrinth is the functional
labyrinth.
Within this system are mem-
bony labyrinth.
located in the petrous portion of the temporal bone,
Chapter 55
Cortical and Brain Stem Control of Motor Function
693
called the
branous tubes and chambers called the membranous
(ductus
semicircular canals; and two large
utricle and
major sensory organ for hearing (see Chapter 52) and
semi-
the
and the saccule are all inte-
gral parts of the equilibrium mechanism.
Located on the inside surface of each utricle and
small sensory area slightly over 2 millimeters in diam-
eter called a macula.
macula of the utricle lies
mainly in the
on the inferior surface
of the utricle and plays an important role in deter-
mining orientation of the head when the head is
macula of the saccule is
located mainly in a vertical plane and signals head
orientation when the person is lying down.
Each macula is covered by a gelatinous layer in
which many small calcium carbonate crystals called
statoconia
sands of
cilia
sensory endings of the
specific gravity two
to three times the specific gravity of the surrounding
the cilia in the direction of gravitational pull.
Directional Sensitivity of the Hair Cells—Kinocilium.
Each
hair cell has 50 to 70 small cilia called stereocilia, plus
Gelatinous
layer
Hair tufts
Nerve fibers
Sustentacular cells
Sustentacular cells
Hair cells
Gelatinous
mass of
cupula
Hair tufts
Utricle
Ampullae
Anterior
Semi-
circular
canals
Maculae and
statoconia
Crista ampullaris
Ductus
cochlearis
Saccule
Posterior
Ductus endolymphaticus
MEMBRANOUS LABYRINTH
Statoconia
CRISTA AMPULLARIS AND MACULA
Nerve
fibers
Hair
cells
Figure 55–9
Membranous labyrinth, and organization of the crista ampullaris
and the macula.
Nerve fiber
Stereocilia
Kinocilium
Filamentous
attachments
Figure 55–10
Hair cell of the equilibrium apparatus and its synapses with the
vestibular nerve.

the pull of gravity. In turn, the vestibular, cerebellar,
terns” of stimulation of the different hair cells apprise
head, different hair cells become stimulated. The “pat-
cles and saccules, so that with different positions of the
the Maintenance of Static Equilibrium
Function of the Utricle and Saccule in
three planes of space.
rate of change
change in rota-
larizes the cells. Then, from the hair cells, appropriate
direction causes depolarization of the hair cells,
tion in the cupula, and bending the cupula in that
hair cells located on the ampullary crest. The kinocilia
causes the cupula to bend to the opposite side.
55–11. Rotation of the head in the opposite direction
strated by the position of the colored cupula in Figure
ampulla, bending the cupula to one side, as demon-
This causes fluid to flow from the duct and through the
direction, the inertia of the fluid in one or more of the
When a person’s head begins to rotate in any
of this crista is a loose gelatinous tissue mass, the
On top
crista ampullaris.
the following manner: Figure 55–11 shows in each
outward.
backward and 45 degrees
outward,
forward and 45 degrees
to the surface of the earth; the anterior ducts are in
bent forward about 30 degrees, the lateral semicircu-
represent all three planes in space. When the head is
lateral (horizontal) semicircular ducts,
anterior, posterior,
vestibular apparatus, known as the
The three semicircular ducts in each
apprises the brain of the head’s orientation in space.
in the gravitational field. It is this “pattern” that
fore, a different pattern of excitation occurs in the
lated when it bends to one side, and so forth. There-
stimulated when it bends backward, others are stimu-
stimulated when the head bends forward, some are
In each macula, each of the hair cells is oriented in
the cilia, appropriate signals are transmitted to the
pletely. Therefore, as the orientation of the head in
decreases the impulse traffic, often turning it off com-
conversely, bending the cilia away from the kinocilium
traffic increases, often to several hundred per second;
stereocilia are bent toward the kinocilium, the impulse
impulses at a rate of about 100 per second. When the
Under normal resting conditions, the nerve fibers
nels, thus causing
tension on the attachments; this closes the ion chan-
versely, bending the pile of stereocilia in the opposite
fluid, causing
numbers of positive ions. Therefore, positive ions pour
opens several hundred fluid channels in the neuronal
eocilia, pulling them outward from the cell body. This
kinocilium bend in the direction of the kinocilium, the
Because of these attachments, when the stereocilia and
next longer stereocilium and, finally, to the kinocilium.
microscope, connect the tip of each stereocilium to the
attachments, almost invisible even to the electron
toward the other side of the cell. Minute filamentous
55–10. The kinocilium is always located to one side,
as shown in Figure
one large cilium, the
The Nervous System: C. Motor and Integrative Neurophysiology
694
Unit XI
kinocilium,
and the stereocilia become progressively shorter
filamentous attachments tug in sequence on the ster-
cell membrane around the bases of the stereocilia,
and these channels are capable of conducting large
into the cell from the surrounding endolymphatic
receptor membrane depolarization. Con-
direction (backward to the kinocilium) reduces the
receptor hyperpolarization.
leading from the hair cells transmit continuous nerve
space changes and the weight of the statoconia bends
brain to control equilibrium.
a different direction so that some of the hair cells are
macular nerve fibers for each orientation of the head
Semicircular Ducts.
and
are
arranged at right angles to one another so that they
lar ducts are approximately horizontal with respect
vertical planes that project
whereas the posterior ducts are in
vertical planes that project
Each semicircular duct has an enlargement at one
of its ends called the ampulla, and the ducts and
ampulla are filled with a fluid called endolymph. Flow
of this fluid through one of the ducts and through its
ampulla excites the sensory organ of the ampulla in
ampulla a small crest called a
cupula.
semicircular ducts causes the fluid to remain station-
ary while the semicircular duct rotates with the head.
Into the cupula are projected hundreds of cilia from
of these hair cells are all oriented in the same direc-
whereas bending it in the opposite direction hyperpo-
signals are sent by way of the vestibular nerve to
apprise the central nervous system of a
tion of the head and the
in each of the
It is especially important that the hair cells are all ori-
ented in different directions in the maculae of the utri-
the brain of the position of the head with respect to
and reticular motor nerve systems of the brain excite
appropriate postural muscles to maintain proper
equilibrium.
Cupula
Ampulla
Cristae
ampullaris
Hair cells
Nerve
rotation.
Figure 55–11
Movement of the cupula and its embedded hairs at the onset of
this has occurred. The semicircular ducts,
ahead of time.
turn to one side,
ducts best by the following illustration: If a person is
We can explain the function of the semicircular
movements.
rapid, intricate changing
ments. Yet loss of function of the semicircular ducts
Therefore, the function of the semicircular ducts is not
to rotate in one direction or another.
All they detect is that the person’s head is
ward direction, one might ask: What is the semicircu-
forward direction, in the side direction, or in the back-
rotating.
to the right in Figure 55–12. Thus, the semicircular
discharge to return to its normal tonic level, as shown
returns to its resting position, thus allowing hair cell
stop discharging entirely. After another few seconds,
bends in the opposite direction, causing the hair cell to
while the semicircular duct stops. This time, the cupula
effects take place: The endolymph continues to rotate
When the rotation suddenly stops, exactly opposite
another 5 to 20 seconds, the cupula slowly returns to
ing as rapidly as the semicircular canal itself; then, in
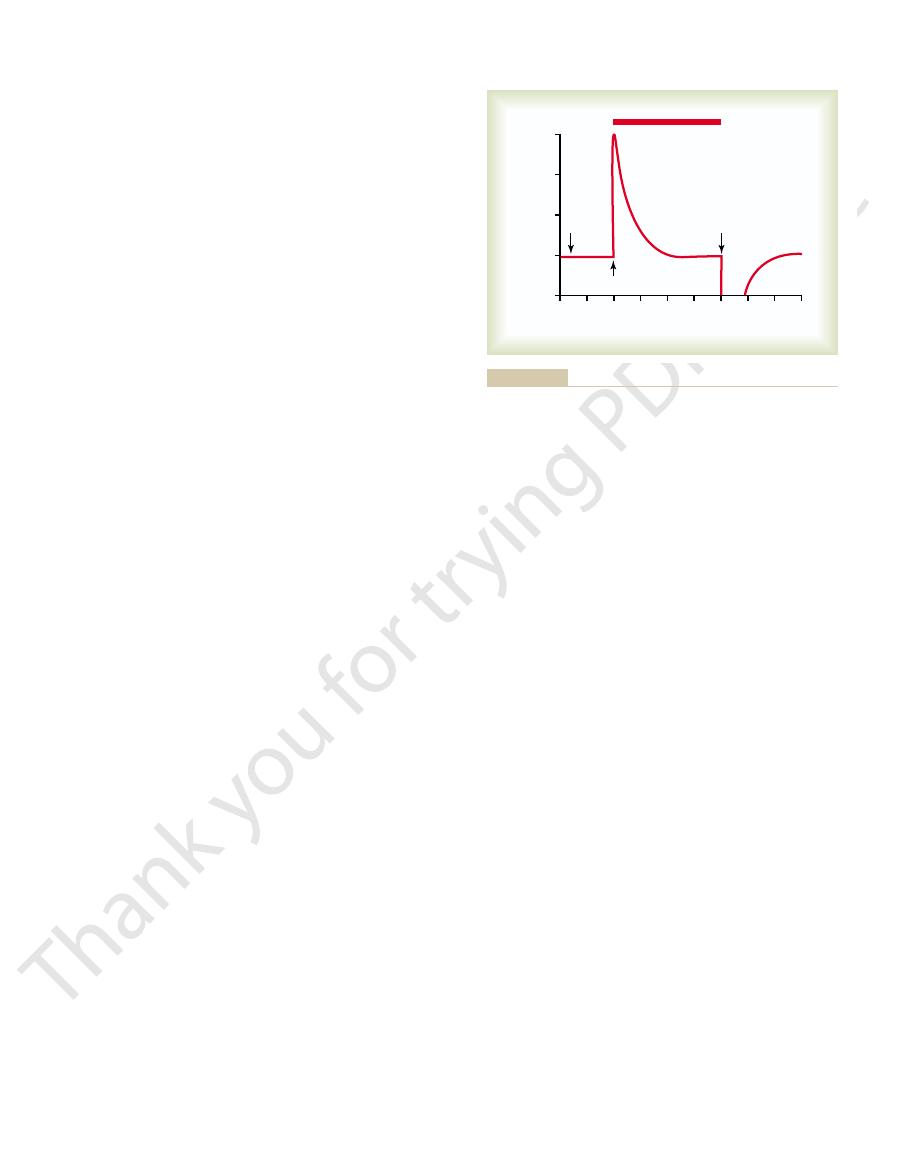
within the first few seconds of rotation, back resistance
The reason for this adaptation of the receptor is that
seconds.
increases greatly; and (3) with continued rotation,
per second; (2) when the animal begins to rotate, the
(1) even when the cupula is in its resting position, the
animal is rotated for 40 seconds, demonstrating that
Figure 55–12 shows a typical discharge signal from
This causes relative fluid flow in the ducts in the direc-
the semicircular ducts, because of its inertia, tends to
), the endolymph in
When the head suddenly begins to rotate in any direc-
Detection of Head Rotation by the
rium adjustments to prevent falling.
organs in the skin, which initiate appropriate equilib-
bodies; in this instance, it is not the maculae that make
running in air, they lean forward to maintain equilib-
vacuum, they would not have to lean forward. When
achieved running speed, if they were running in a
When runners first begin to run, they
velocity.
The maculae
Thus, the maculae operate to maintain equilibrium
equilibrium and leans the body forward no farther.
this point, the nervous system senses a state of proper
conia to fall backward because of the acceleration. At
statoconia exactly equals the tendency for the stato-
falling backward. This automatically causes the person
tion of dysequilibrium is sent into the nervous centers,
fluid, fall backward on the hair cell cilia, and informa-
that is, when the body accelerates—the statoconia,
When the body is suddenly thrust forward—
is in the near-vertical position. Indeed, a person
This utricle and saccule system functions extremely
Chapter 55
Cortical and Brain Stem Control of Motor Function
695
effectively for maintaining equilibrium when the head
can determine as little as half a degree of dysequilib-
rium when the body leans from the precise upright
position.
Detection of Linear Acceleration by the Utricle and Saccule
Maculae.
which have greater mass inertia than the surrounding
causing the person to feel as though he or she were
to lean forward until the resulting anterior shift of the
during linear acceleration in exactly the same manner
as they operate during static equilibrium.
do not operate for the detection of
linear
must lean far forward to keep from falling backward
because of initial acceleration, but once they have
rium only because of air resistance against their
them lean but air pressure acting on pressure end-
Semicircular Ducts
tion (called angular acceleration
remain stationary while the semicircular ducts turn.
tion opposite to head rotation.
a single hair cell in the crista ampullaris when an
hair cell emits a tonic discharge of about 100 impulses
hairs bend to one side and the rate of discharge
the excess discharge of the hair cell gradually
subsides back to the resting level during the next few
to the flow of fluid in the semicircular duct and past
the bent cupula causes the endolymph to begin rotat-
its resting position in the middle of the ampulla
because of its own elastic recoil.
the endolymph stops moving and the cupula gradually
duct transmits a signal of one polarity when the head
begins to rotate and of opposite polarity when it stops
“Predictive” Function of the Semicircular Duct System in the
Maintenance of Equilibrium.
Because the semicircular
ducts do not detect that the body is off balance in the
lar ducts’ function in the maintenance of equilibrium?
beginning
or stopping
to maintain static equilibrium or to maintain equilib-
rium during steady directional or rotational move-
does cause a person to have poor equilibrium when
attempting to perform
body
running forward rapidly and then suddenly begins to
he or she will fall off balance a frac-
tion of a second later unless appropriate corrections
are made
But the maculae of the utricle
and saccule cannot detect that he or she is off balance
until after
0
10
20
30
40
50
60
70
80
90
Impulses per second
0
100
200
300
400
Rotation
Stop rotation
Begin rotation
Tonic
level of
discharge
Seconds
Seconds
first by the onset of head rotation and then by stopping rotation.
Response of a hair cell when a semicircular canal is stimulated
Figure 55–12

lospinal tracts, the medial longitudinal fasciculus, and
that also send fibers into the cerebellum, the vestibu-
lobe nuclei. The fibers that end in the brain stem
to the cerebellar fastigial, uvular, and flocculonodular
medulla and the pons. Some fibers pass directly to the
the vestibular nerve. Most of the vestibular nerve fibers
Figure 55–13 shows the connections in the hindbrain of
Central Nervous System
Neuronal Connections of the Vestibular Apparatus with the
eyes are closed, equilibrium is immediately lost.
formed slowly. But when moving rapidly or when the
centers. Some people with bilateral destruction of the
the body, a person can still use the visual mechanisms
After destruction of the vestibular apparatus, and
result, the person leans forward to oppose this.
different from that caused by gravitational pull; as a
running. The air pressure against the front of the body
maintenance of equilibrium. For instance, pressure
of the Body.
the entire body.
the neck proprioceptors; therefore, in this case, the
leans in one direction, the impulses from the
ted from the vestibular apparatus. However,
person a sense of dysequilibrium. They do this by trans-
When the
joint receptors of the neck.
head with respect to the body. This information is trans-
Therefore,
The vestibular apparatus detects the
with Equilibrium
These reflexes are described
head. This results from reflexes transmitted through the
nately, each time the head is suddenly rotated, signals
each object long enough to gain a clear image. Fortu-
detecting an image unless they remained “fixed” on
eyes’ gaze. In addition, the eyes would be of little use in
or backward, it would be impossible to maintain a stable
ment rapidly or even leans the head sideways, forward,
When a person changes his or her direction of move-
Vestibular Mechanisms for
discussed in the following chapter.
librium. These other functions of the cerebellum are
the body, as well as for those having to do with equi-
as a “predictive” organ for most rapid movements of
signals but has less effect on detecting macular signals.
priate anticipatory preventive adjustments. In this way,
In other words, the semicircular duct mechanism
is made.
turning, and this information can easily apprise the
however, will have already detected that the person is
The Nervous System: C. Motor and Integrative Neurophysiology
696
Unit XI
central nervous system of the fact that the person will
fall off balance within the next fraction of a second or
so unless some anticipatory correction
predicts that dysequilibrium is going to occur and
thereby causes the equilibrium centers to make appro-
the person need not fall off balance at all before he or
she begins to correct the situation.
Removal of the flocculonodular lobes of the cere-
bellum prevents normal detection of semicircular duct
It is especially interesting that the cerebellum serves
Stabilizing the Eyes
image on the retinas unless the person had some auto-
matic control mechanism to stabilize the direction of the
from the semicircular ducts cause the eyes to rotate in
a direction equal and opposite to the rotation of the
vestibular nuclei and the medial longitudinal fasciculus
to the oculomotor nuclei.
in Chapter 51.
Other Factors Concerned
Neck Proprioceptors.
orientation and movement only of the head.
it is essential that the nervous centers also receive
appropriate information about the orientation of the
mitted from the proprioceptors of the neck and body
directly to the vestibular and reticular nuclei in the
brain stem and indirectly by way of the cerebellum.
Among the most important proprioceptive informa-
tion needed for the maintenance of equilibrium is that
transmitted by
head is leaned in one direction by bending the neck,
impulses from the neck proprioceptors keep the signals
originating in the vestibular apparatus from giving the
mitting signals that exactly oppose the signals transmit-
when the
entire body
vestibular apparatus are not opposed by signals from
person does perceive a change in equilibrium status of
Proprioceptive and Exteroceptive Information from Other Parts
Proprioceptive information from parts of
the body other than the neck is also important in the
sensations from the footpads tell one (1) whether
weight is distributed equally between the two feet and
(2) whether weight on the feet is more forward or
backward.
Exteroceptive information is especially necessary for
the maintenance of equilibrium when a person is
signals that a force is opposing the body in a direction
Importance of Visual Information in the Maintenance of Equilib-
rium.
even after loss of most proprioceptive information from
reasonably effectively for maintaining equilibrium.
Even a slight linear or rotational movement of the body
instantaneously shifts the visual images on the retina,
and this information is relayed to the equilibrium
vestibular apparatus have almost normal equilibrium as
long as their eyes are open and all motions are per-
terminate in the brain stem in the vestibular nuclei,
which are located approximately at the junction of the
brain stem reticular nuclei without synapsing and also
vestibular nuclei synapse with second-order neurons
Recticulospinal
tract
Rubrospinal tract
Vestibulospinal tract
Fastigioreticular
tract
Reticular
substance
Red
nucleus
Medial longitudinal
fasciculus
Fastigial
nucleus
Dentate nucleus
Vestibular nucleus
Flocculonodular
lobe
Vestibular nerve
large oval white area) with other areas of the central nervous
Connections of vestibular nerves through the vestibular nuclei (the
Figure 55–13
system.

47:395, 1998.
macology of motion sickness: an update. Brain Res Bull
Yates BJ, Miller AD, Lucot JB: Physiological basis and phar-
cal motor system. Curr Biol 14:R204, 2004.
Umilta MA: Frontal cortex: goal-relatedness and the corti-
non-human primates. Curr Opin Neurobiol 13:671, 2003.
movements: insights from neurophysiological studies on
Scott SH: The role of primary motor cortex in goal-directed
Curr Biol 14:R353, 2004.
Schieber MH: Motor control: basic units of cortical output?
Opin Neurobiol 13:225, 2003.
Sanes JN: Neocortical mechanisms in motor learning. Curr
during motor action. Curr Opin Neurobiol 13:678, 2003.
Salenius S, Hari R: Synchronous cortical oscillatory activity
cochlea. Physiol Rev 81:1305, 2001.
Robles L, Ruggero MA: Mechanics of the mammalian
cochlea. Brain Res Bull 60:397, 2003.
Raphael Y, Altschuler RA: Structure and innervation of the
incomplete spinal cord injury. Nat Rev Neurosci 2:263,
Raineteau O, Schwab ME: Plasticity of motor systems after
Curr Biol 13:R802, 2003.
Johansen-Berg H: Motor physiology: a brain of two halves.
Brain Res Brain Res Rev 40:152, 2002.
Garwicz M: Spinal reflexes provide motor error signals to
Neurophysiol 91:1919, 2004.
system during active versus passive head movements. J
Cullen KE, Roy JE: Signal processing in the vestibular
head movement. Ann N Y Acad Sci 942:364, 2001.
Boyle R: Vestibulospinal control of reflex and voluntary
hand in the cerebral cortex. Behav Brain Res 135:179,
Blake DT, Byl NN, Merzenich MM: Representation of the
and posterior cerebellum. Brain Res Bull 60:511, 2003.
Barmack NH: Central vestibular system: vestibular nuclei
Vestib Res 13:245, 2003.
processing of vestibular signals to detect motion or tilt. J
Angelaki DE, Dickman JD: Gravity or translation: central
processing. Curr Opin Neurobiol 13:440, 2003.
Alitto HJ, Usrey WM: Corticothalamic feedback and sensory
causes them to pull to the sitting position. It is clear that
objects with movements of the eyes and head. Also,
can yawn and stretch. They can cry and can follow
hands to the mouth to suck the fingers. In addition, they
sion of unpleasant food from the mouth, and moving the
typed movements for feeding, such as suckling, extru-
many months. They are able to perform some stereo-
cephaly.
the mesencephalic region, a condition called
Rarely, a baby is born without brain structures above
Subconscious, Stereotyped
Nuclei in Controlling
apprise the psyche of the equilibrium status of the body.
area of the superior temporal gyrus. These signals
or through reticular tracts) to the cerebral cortex, ter-
movements of the eyes every time the head rotates, so
rium under static conditions. It is believed that the
rapid changes in direc-
destruction of the semicircular ducts themselves. That is,
the semicircular ducts. In fact, destruction of these lobes
The
inhibition of the many antigravity muscles, thus auto-
vestibulospinal and reticulospinal tracts. The signals to
stem, as well as down the spinal cord by way of the
passes to the vestibular nuclei and cerebellum. Next,
excited by the vestibular apparatus. The pathway then
begins in the vestibular nerves, where the nerves are
The primary pathway for the equilibrium reflexes
other areas of the brain stem, particularly the reticular
Chapter 55
Cortical and Brain Stem Control of Motor Function
697
nuclei.
signals are sent into the reticular nuclei of the brain
the cord control the interplay between facilitation and
matically controlling equilibrium.
flocculonodular lobes of the cerebellum are espe-
cially concerned with dynamic equilibrium signals from
results in almost exactly the same clinical symptoms as
severe injury to either the lobes or the ducts causes loss
of dynamic equilibrium during
tion of motion but does not seriously disturb equilib-
uvula
of the cerebellum plays a similar important role in static
equilibrium.
Signals transmitted upward in the brain stem from
both the vestibular nuclei and the cerebellum by way of
the medial longitudinal fasciculus cause corrective
that the eyes remain fixed on a specific visual object.
Signals also pass upward (either through this same tract
minating in a primary cortical center for equilibrium
located in the parietal lobe deep in the sylvian fissure
on the opposite side of the fissure from the auditory
Functions of Brain Stem
Movements
anen-
Some of these babies have been kept alive for
placing pressure on the upper anterior parts of their legs
many of the stereotyped motor functions of the human
being are integrated in the brain stem.
References
2002.
cerebellar modules—relevance for motor coordination.
2001.