Bioinformatics المعلوماتية الحيوية
/ المرحلة الكورس الثاني / فيزياء حياتية
الدكتور عبدالرحيم ذنون الغزال
المصدر
كتاب /المعلوماتية الحيوية
دكتورة زهرة محمود الخفاجي والدكتور علي عبد الحافظ إبراهيم
معهد الهندسة الوراثية والتقنيات الحيوية قسم التقنيات الحيوية \ كلية العلوم \
للدراسات العليا \ جامعة بغداد جامعة النهرين \ العراق
2012
1
الطفرات
على مستوى ضيق او مدى واسع وذلك لان الجينات وحدات قابلة DNA تغير في توالياتللتطفير وقد أصبحت الحجر الأساس في الدراسات الوراثية المبكرة وهناك بعض الحقائق العامة
حول الطفرات ومنها:
في الأحياء الراقية تؤدي الى التطور. Germ cells الطفرات في ·
انواع الطفرات حسب موقعها:-1-طفرات تؤثر في جين واحد كما في طفرات الخلايا الحقيقية النواة المؤثرة في وحدة ·
انتساخ واحدة وأخرى تؤثر في اثنين من الجينات ذات العلاقة وأخرى تؤثر في عدة جينات
خاصة اذا كانت في التواليات المنظمة كما في الخلايا بدائية النواة عندما تقع Pleiotropic
الجينات في اوبرون واحد.
2-الطفرات في الاكسونات او الانترونات تؤدي في بعض الأحيان الى الأمراض الوراثية. ·
حدوث الطفرات يمكن ان يؤثر في عمليات التضاعف والانتساخ . ·
3-الطفرات الواقعة في المناطق غير المشفرة يمكن ان يكون لها تأثيرا" كبيرا" فمثلا في الخلايا ·
حقيقية النواة تسيطر بعض هذه المناطق في تخصيص مواقع فلق عند النهاية ' 3 او إضافة ذيل الأدنين
وحصول الطفرات في هذه mRNA او عمليات خياطة النسخ الأولية من Poly (A) site
mRNA وإنتاج (Heterogeneous nuclear RNA) hn RNA المناطق تمنع معالجة
الناضج الفعال وبالتالي تمنع التشفير للبروتينات.
2
هل الجينوم وحدة ثابتة؟
تراكم الطفرات في الجينوم على مرور الزمن يغير من الجينوم وعليه فان الجينوم ليس ·وحدة ثابتة وانما يتغير عبر الزمن أثناء عملية التطور
بعض الطفرات تسبب الأمراض كما في حدوث السرطانات ·
بعض الطفرات قد تكون حميدة وتزيد من تلاؤم الكائن مع بيئته وأخرى قد تكون كارثية ·
تؤدي الى موت الكائن، والأخيرة تحدث عندما يتغير بروتين مهم للخلايا ويصبح غير فعال
مما يؤدي الى موت الخلايا وفي أحيان أخرى قد تكون بدون تأثير محسوس.
وعندما تكون
الطفرة مفيدة وتساعد الكائن لابد ان تتوارث في الأجيال الناتجة خلال عملية التكاثر وإلاسيذهب تأثيرها بانتهاء الفرد كما ذكر أعلاه.
3
أنواع الطفرات
هناك عدة أسس لتقسيم الطفرات منها حسب النمط المظهري:النمط المظهري الذي ينتج عن حدوث الطفرة وفي هذا المجال يمكن ذكر
وهذه يمكن ان تحدث في أ- المناطق الموجودة بين Silent mutants 1-الطفرات الصامتة ·
الجينات او في ب - المناطق غير المشفرة في الجين، او تحدث ج - في المناطق المشفرة دون
التأثير في البروتينات الناتجة وهذا يحصل بالنسبة للحوامض الامينية التي لها أكثر من
شفرة مترادفة، اذ ان التغير سيؤدي الى إنتاج الحامض الاميني نفسه كما موضح في
. ( الشكل ( 24.3
وهذا يكون وراء ظهور السلالات المتطابقة من حيث المواصفات وتوالي الحوامض الامينية
في بروتيناتها ولكنها تظهر الاختلاف في توالي جينوماتها
شكل 24.3 : الطفرات الصامتة
4
Point طفرات تكون موضعية او نقطية ،Missense mutants 2- الطفرات الخاطئة ·
اي يحصل تغير في قاعدة نتروجينية واحدة يؤدي الى تغير الحامض mutation
الاميني المشفر له وبالتالي يدمج حامض اميني غير صحيح ويؤدي الى تغير البروتين
وفي اغلب الأحيان تكون هذه الطفرات يمكن تحملها اذ لا تؤدي الى تغيير كبير في وظيفة
البروتين وفي بعض الأحيان يكون لها تأثير كبير
شكل 25.3 : الطفرات الخاطئة
.
.
5
طفرات تحدث بحيث تغير الشفرة الوراثية Nonsense mutants 3-الطفرات التافهة ·
للحوامض الامينية الى شفرة وقف تؤدي الى إنهاء عملية الترجمة مما يؤدي الى تقصير
. طول السلسلة الببتيدية الناتجة من الجين كما موضح في الشكل ( 26.3
وتأثيرها يعتمد على الموقع الذي حصلت فيه ولكن في العادة تكون كارثية وتنتج بروتين مبتور
غير فعال
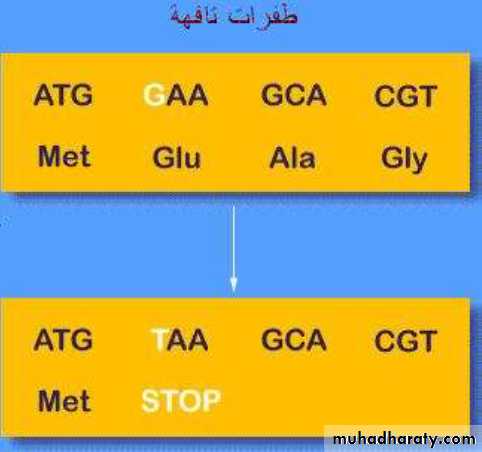
شكل 26.3 : الطفرات التافهة
6طفرات نقطية ايضا تؤدي :Readthrough mutants 4- طفرات القراءة المستمرة ·
الى تغير شفرة الوقف الى شفرة لأحد الحوامض الامينية مما يؤدي الى امتداد البروتين
المتعددة. ومعظم mRNA الى ان يصل الى شفرة وقف قادمة خاصة في رسائل
البروتينات تتحمل الاستطالة البسيطة دون التأثير في وظيفتها ولكن الاستطالات الكبيرة
تؤثر في طوي البروتين وفي العادة تؤدي الى تخفيض فعاليته.
طفرات :Frameshift reading mutants 5- طفرات إزاحة أطر القراءة الصحيحة ·
نقطية بطبيعتها وتكون اما إضافة او إزالة قاعدة نتروجينية واحدة في التواليات المشفرة
مما يؤدي الى تغير كل الشفرات الوراثية التي تقع الى يمينها كما موضح في الشكل
: 27.3
7
اذ ان إزالة قاعدة واحدة سيزحف تأثيرها في الشفرات التي بعدها وتتغير الحوامض الامينية في
السلاسل الببتيدية الناتجة
.
6- طفرات في مواقع الخياطة: طفرات تحدث في المنطقة الحدودية بين الانترونات ·
والاكسونات في جينومات الخلايا حقيقية النواة تؤدي الى اضطراب خياطة الجين
والطفرات يمكن ان تؤدي الى عدم إمكانية إزالة الانترونات من
ولكن الخلايا يمكن ان تستعمل مواقع بديلة وهي المواقع RNA النسخ الأولية لل
الخفية وتتمثل بتواليات تشبه التواليات الأصلية وربما نتجت من حدوث طفرات مثل
طفرات إزاحة أطر القراءة وبعض الأحيان يكون وجود التواليات الخفية أفضل من
الأصلية .
ان حدوث اضطرابات في مواقع الخياطة يؤدي اما الى إضافة جزء من البروتين او حذف جزء
.Thalassemia منه (للجين تحت التعبير) وفي العادة ترتبط ببعض الأمراض مثل الثلاسيميا
8
العوامل المطفرة
هناك عددا" من المواد الكيماوية او العوامل الفيزياوية الموجودة في البيئة المحيطة بالخلايا تحث
الطفرات وبذا فهي تمثل الرابط بين الوراثة والبيئة، إضافة الى ان الطفرات يمكن ان تحصل
بشكل تلقائي. وتختلف العوامل المطفرة في تأثيرها أثناء عملية التطفير.
وفي العموم فان المطفرات يمكن ان تؤدي الى
1-الأكسدة او
2-حث المثيلة غير المسيطر عليها او
3-التحلل او
4-تحوير القواعد النتروجينية ، وفي التالي بعض الحالات :
بعض المواد الفعالة
5- تغير تركيب القواعد النتروجينية الى تراكيب كيماوية مختلفة او تؤدي ·
الى إزالة القواعد النتروجينية.
9
فكل من القواعد النتروجينية مثل السايتوزين والأدنين والكوانين تحوي على مجموعة أمين التي
ومن العوامل التي لها DNA يمكن ان تزال مؤدية الى عدم انتظام تكامل القواعد في أشرطة
ومن جهة ثانية فان إزالة مجموعة الأمين يمكن . Nitrous acid هذا التأثير حامض النتروز
ان يحدث بشكل تلقائي بمعدل 100 تغيير قاعدة \ الجينوم \ يوم ، وأكثر المناطق التي تحدث
فيها هذه العملية هي قاعدة السايتوسين في الموقع 4 لذا تعد هذه بمثابة مناطق ساخنة، وينتج
من عملية إزالة الأمين تكوين القاعدة النتروجينية اليوراسيل التي ترتبط مع الأدنين بدلا من
غير مسموح به، لذا تقوم الخلايا بإصلاح هذا DNA الكوانين، ووجود اليوراسيل في أشرطة
وربما بآليات أخرى. Uracil-DNA glycosylase العطب بكفاءة بواسطة الإنزيم
10
اما التفاعل الآخر الذي يحصل في البقع الساخنة فهي إضافة مجموعة مثيل على الذرة الخامسة
Methyl cytosine للسايتوزين وإنتاج
وهناك تحويرات أخرى .
يمكن ان تطول القواعد النتروجينية الأخرى .
بعض المواد يمكن ان تؤدي الى إزالة قاعدة نتروجينية او امتداد من النيوكلوتيدات وهذا ما ·
يسمى بطفرات الشطب او الحذف ، والامتدادات المحذوفة قد تكون جين واحد او عدد من
.Clastogenic agent الجينات وعندما تكون الامتدادات كبيرة تسمى العوامل
11
وفي حالة التأثير على مستوى القواعد النتروجينية فهذه قد تكون إزالة شفرة واحدة او قاعدة نتروجينية
واحدة من الشفرة . ومن أفضل الأمثلة عليها إزالة البيورينات وقد تم تقدير ان هناك حوالي
5000 قاعدة بيورينية اما أدنين او كوانين يمكن ان تزال من الجينوم \ يوم ويكون ذلك نتيجة
للسكر منقوص N-glycosyl linkage لتأثير الحرارة. وتتم اما بكسر الاصرة
الأوكسجين مع نتروجين الكوانين في الموقع 9 وينتج عنه سكر بدون بيورين وبدوره يؤدي
الى عملية الحذف في الدورة القادمة من التضاعف ان لم يتم إصلاحها كما موضح في الشكل
:28.3
شكل 28.3 : حدوث طفرات الحذف
12
وتعني إضافة مجموعة مثيل او اثيل الى القواعد النتروجينية، Alkylation عملية الألكلة ·
فألكلة الكوانين مثلا تشمل إحلال آصرة الهيدروجين لذرة الأوكسجين في الموقع السادس
6- التي لا يمكن ان ترتبط بالسايتوزين وإنما methyl guanine بمجموعة مثيل لتكوين
ترتبط بالثايمين وعليه وبعد دورة تضاعف قادمة فان السايتوزين المقابلة يتم استبدالها
بالثايمين في الجزيئة الجديدة كما موضح في الشكل 29.3
شكل 29.3 : ألكلة القواعد النتروجينية
Ethyl-nitroso-urea (ENU) ومن العوامل المسببة لهذه الحالة
Ethyl methane-sulfonate (EMS)
Dimethyl-nitros-amine
N-methyl-N-nitro-N' Nitroso-guanidine (NTG)
13
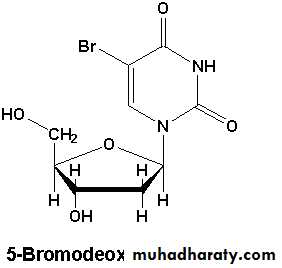
Nucleotide base analogue متشابهات قواعد النيوكلوتيدات
نظرا لتشابهها مع قواعده وتستطيع ان تندمج بالأشرطة أثناء DNA وهي مركبات تؤدي الى تطفير5) تشابه الثايمين وتحوي على ذرة BrdU) 5-Bromo-deoxy-uridine تضاعفها. فالمادة
بروم بدلا من مجموعة المثيل في الموقع 5 وبذا تستطيع ان تندمج أثناء التضاعف ، ووجود
ويؤدي الى اضطراب تكامل القواعد وبالتالي الخطأ DNA ذرة البروم يكون غير ملائما في
في ازدواجها.
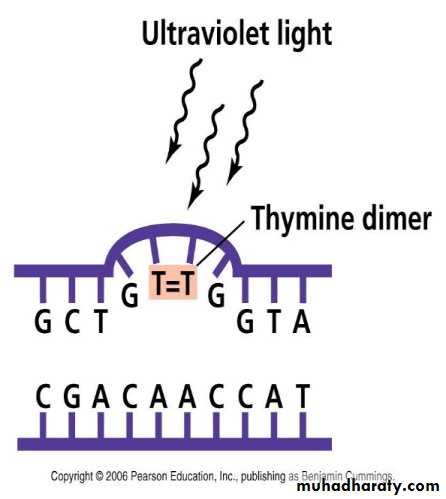
الأشعة فوق البنفسجية
الأشعة عند الأطوال الموجية القصيرة منها مثل 260 نانومتر وبالتحديد 254 نانومتر التي تمثل
للأشعة تؤدي الى تكوين أواصر تساهمية بين ثمالات الثايمين DNA ذروة امتصاص
مكونة مزدوجات الثايمين ويؤدي تكوينها الى التداخل السلبي مع ، المتجاورة عند الموقع 5
14
الطفرات التلقائية
الطفرات التي تحدث دون الحث بالمواد المطفرة ويمكن ان تنتج عن:فالمعروف ان تضاعف الأشرطة يكون بشكل شبه ، DNA أخطاء أثناء عملية تضاعف ·
محافظ ويعتمد على أساس تكامل القواعد في الأشرطة ، ويمكن ان يحدث الخطأ أثناءالتضاعف بمعدل
1 في 510 قاعدة وينخفض التردد الى 1 في 710
وإذا لم تصحح الطفرة فإنها تعبر الى الخلايا الناتجة ويكون تأثيرها معتمدا على مكان
وجودها مثلا في التواليات المشفرة فانه سيؤثر في وظيفة البروتين الناتج.
.
15
Replication slippage طفرات تحدث نتيجة لانزلاق او تفاوت عملية انزلاق التضاعف ·
وهذه بطبيعة الحال لا تندرج تحت المجاميع المدرجة أعلاه ، وتنتج من عدم الاصطفاف
الاليلية وغير الاليلية أثناء عمليات التضاعف فمثلا عند احتواء DNA المضبوط بين تواليات
كما في التوابع الدقيقة التوابع CA repeats أشرطة القالب على مكررات من القواعد مثل
الدقيقة ، فان الأشرطة الجديدة والقوالب يمكن ان يحصل زحف
Polymerase في مواقعها واحدا بالنسبة للأخرى وبالتضاعف يحصل انزلاق لأنزيم الكوثرة
مما يؤدي الى أخطاء في ازدواج المكررات حيث تنتسخ بعض المكررات مرتين slippage
في حين لا تنتسخ الأخرى اعتمادا على اتجاه الزحف الحاصل
16
البروتينات
تمثل النواتج النهائية لعمليات التعبير الجيني (الانتساخ) والتعبير ألبروتيني (الترجمة) ،
البروتينات مكوثرات حيوية هي الأكثر تنوعا ، وحدات بناءها الحوامض الامينية، وتعد من
-1) Nanoparticles الجزيئات الحيوية العملاقة ولكنها تبقى ضمن مدى الجزيئات النانوية
100 نانومتر) اعتمادا على حجمها الفيزيائي، وتسمى حصان الشغل لأنها مسئولة عن وظائف
متعددة مثل الوظائف التركيبية والنقل والتنظيم وتضخيم الإشارات وغيرها.
تركيبها الأساسي (الأولي) يتكون من سلسلة من الحوامض الامينية التي ترتبط بآصرة ببتيدية
بعد ارتباطها مع بعضها، والجزيئات المكونة من 40 Residue التي تعرف كل منها بالثمالة
ثمالة يطلق عليها الببتيدات .،
17
وتحوي البروتينات العديد من المجاميع الفعالة مثل 1-الكحولات
فضلا Carboxamide 3- واسترات الثايول 4-ومجاميع الكربوكسيل 5-ومجاميع Thiols 2-والثايولات
عن وجود 6-المجاميع القاعدية، وهذه المجاميع يمكن ان تحدد الأطر العامة لفعالية البروتينات.
والمجموع البروتيني الكلي لاي خلية يدعى بمجال المعلوماتية الحيوية ب المكنون ألبروتيني ويعني كل
مجاميع البروتينات المعبر عنها في الخلية اي انها المنتجات المترجمة للجينوم ، و
يعنى دراسة كل من 1- المكنون ألبروتيني 2- دراسة كل التحليلات المتزامنة Proteomics
للبروتينات المترجمة في الخلية، 3- تحديد كميات البروتينات ومواقعها في الخلية
4-دراسة كل التحويرات بالبروتينات التي تجري عليها بعد الترجمة والتداخلات فيما بينها ، 5-تحديد وظائف البروتينات،
ومثل هذه الدراسات تؤدي الى معرفة وفهم الوظائف الخلوية للبروتينات وبالتالي إدراك وظائف
الجينات
18
تقوم البروتينات بكل الوظائف الحيوية (تقريبا) التي تجري في الأنظمة الحيوية وبصفة رئيسة
كإنزيمات ، ولكن هذه الفعاليات تتم بعد ان تجري العديد من التحويرات على الجزيئات الأولية
التي تتم بعدد من التداخلات مثل 1-الأواصر الهيدروجينية Folding التي تعاني من الطوي
2-والتداخلات الكارهة للماء وغيرها من التداخلات .
3-المشاركة الفاعلة في عمليات انقسام ونمو الخلايا والأنسجة وتمايزها، 4-والسيطرة
على نقل الإشارات من خارج الأنظمة الحيوية الى داخلها،
5-وكذلك السيطرة على التوازن
ألايوني للجسم بالسيطرة على نقل المواد من والى الخلايا،
6-وتساهم ايضا في الوظائف
الميكانيكية كما في الجلد والأوعية الدموية وغيرها من المناطق
7-مساهمتها في بناء
المنصات والهياكل الخلوية وكذلك في الأنسجة الرابطة والأنسجة الواقية
19
وتكمن أهمية دراسة البروتينات في كونها تمثل الناتج المباشر للتعبير عن المعلومات الوراثية.
فضلا عن انها عصب الحياة في التفاعلات التي تجري وان الجزيئات الحيوية الأخرى تكاد
تكون نواتج لفعالياتها.
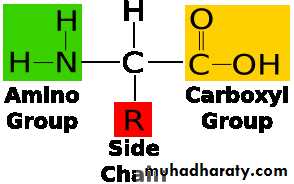
الحوامض الامينية
ترتبط بأربعة مجاميع، ابسطها Ca جزيئات حيوية تركيبها الأساسي ذرة كربون يطلق عليها
(-COOH) وكذلك مجموعة الكربوكسيل (-NH الهيدروجين، وترتبط الى مجموعة ألامين ( 2
والتي تكون R وهذه المجاميع عادة تكون ثابتة، اما الموقع الرابع فيرتبط الى السلسلة الجانبية
مختلفة وهي التي تضفي الصفات على الحامض الاميني، والتركيب الأساسي موضح في الشكل
: 69.3
شكل 69.3 : تركيب الحامض الاميني
20
ويرمز للحوامض الامينية اما بثلاث حرف الاولى للحامض
V او بحرف واحد Val تكون Valine
21
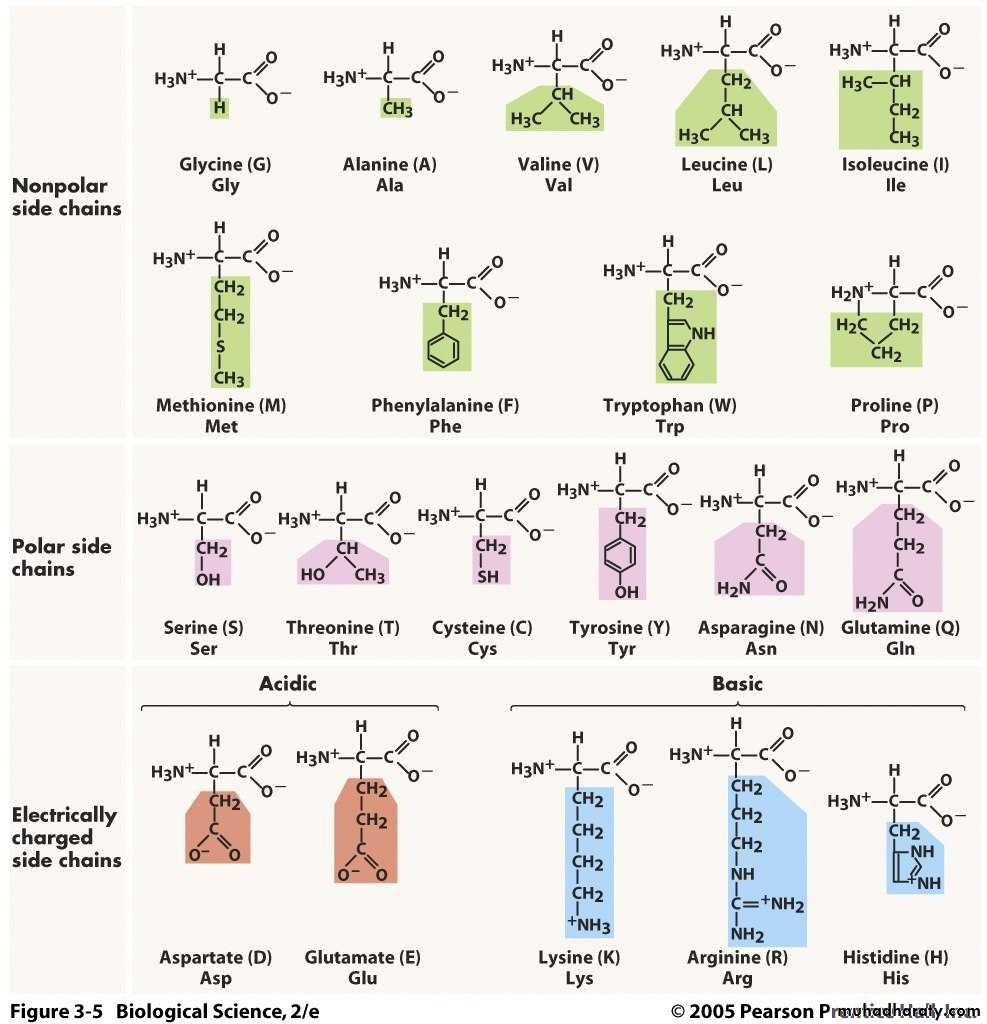
Hydrophobic
Hydrophilic
Negative charged A.A Positive A.A-
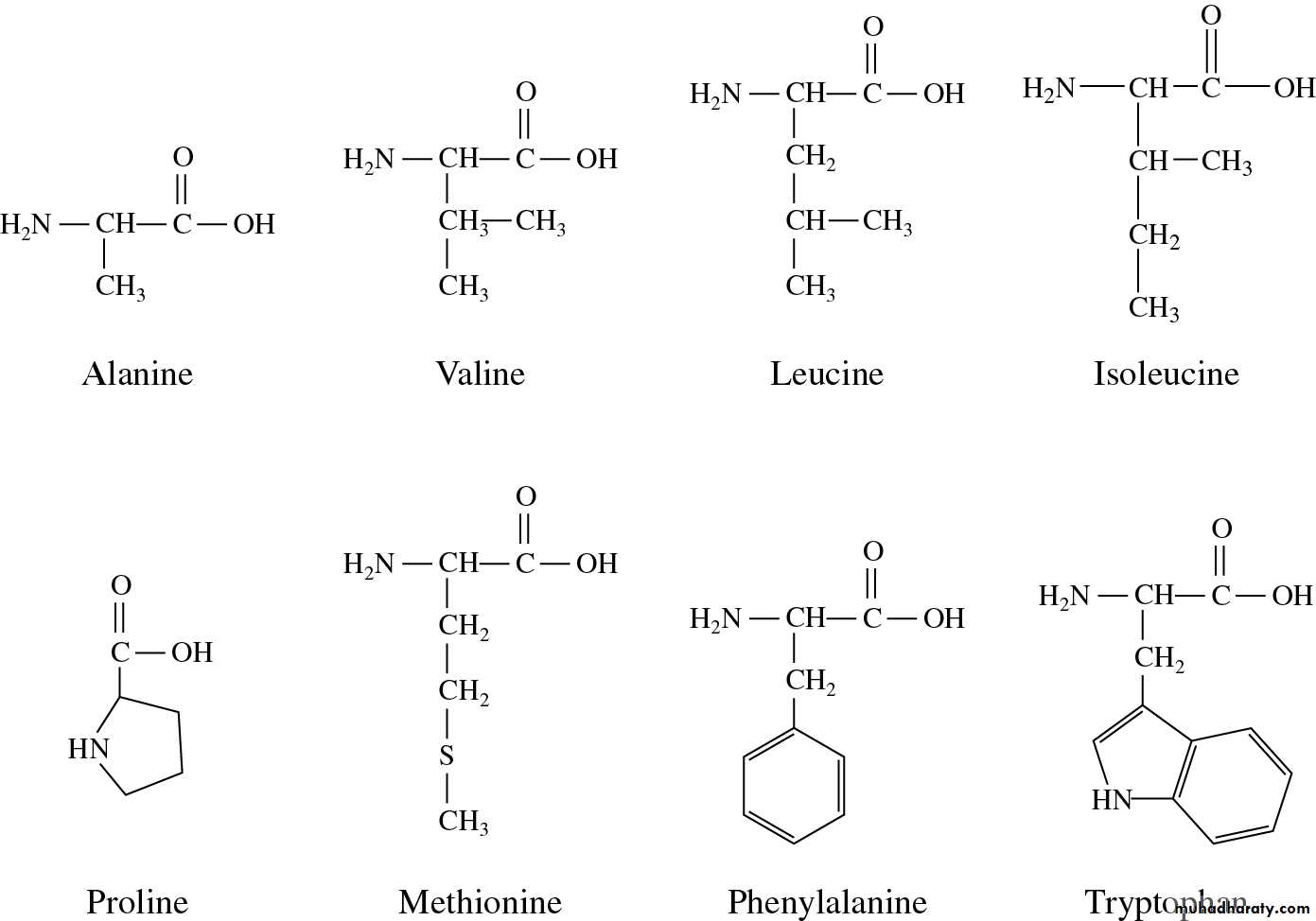
Hydrophobic الحوامض الامينية الكارهة للماء
تشمل الليوسين والايزوليوسين والفنيل-النين والفالين، وبدرجة اقل التايروسين ، الالنين
والتربتوفان والسستئين . وهذه تقسم بدورها الى حوامض أمينية كارهة للماء اليفاتية
مكونة من سلسلة كربونية مستقيمة مثل الالنين والفالين والليوسين والايزوليوسين Aliphatic
والميثايونين، في حين تكون المجموعة الثانية هي الحلقية والتي تحوي على حلقات مختلفة مثل
الفنيل-النين والتربتوفان ويلحق بها البرولين.
.
22
.
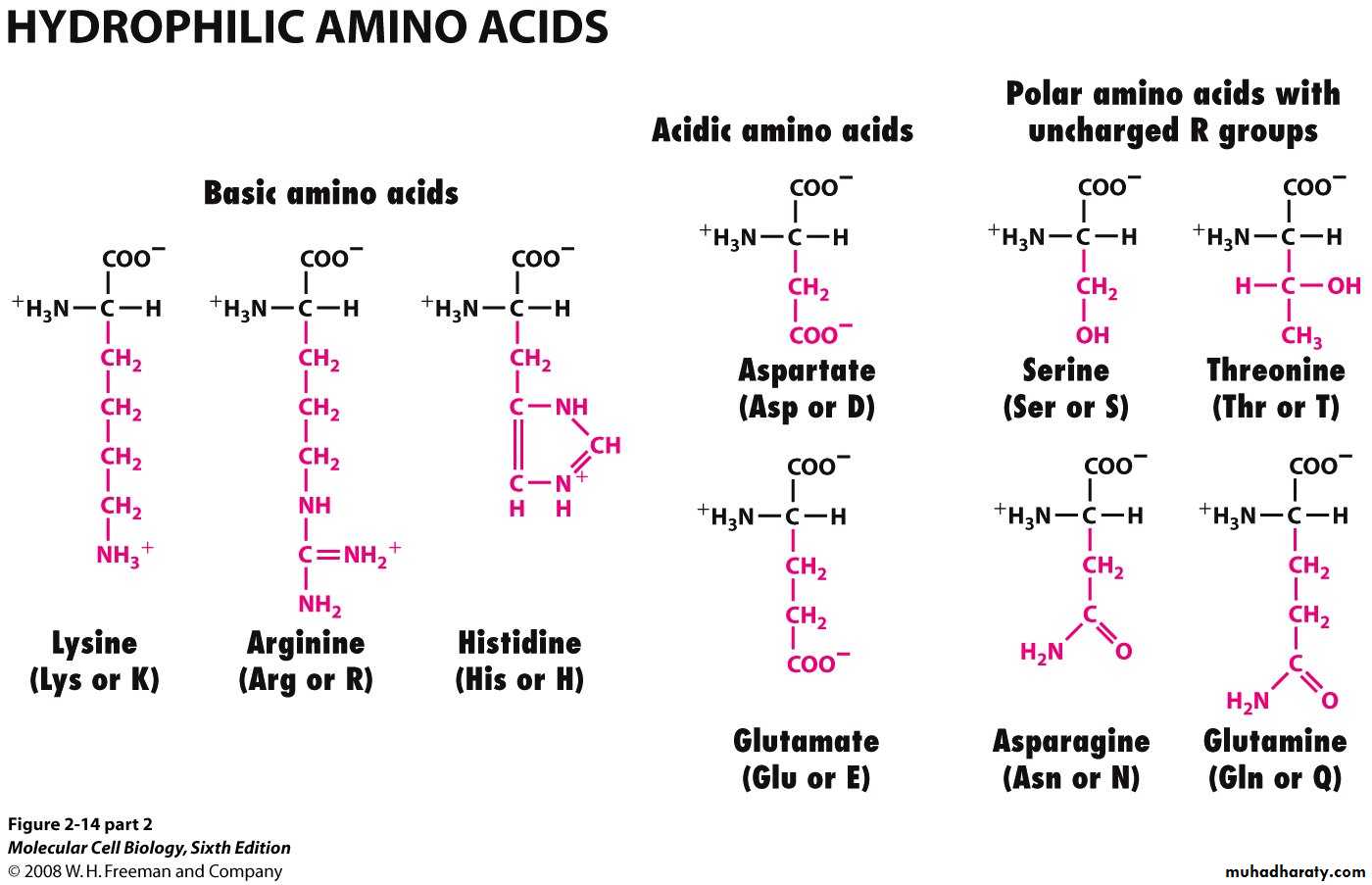
Hydrophilic الحوامض الامينية المحبة للماء
تضم السيرين والثريونين التايروسين والاسبارجين والكلوتامين. وقد تكون هذه المجموعة قطبية
او غير قطبية
23
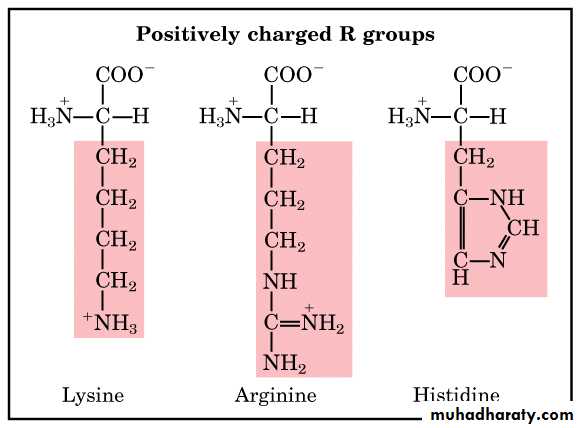
الحوامض الامينية المشحونة بشحنة موجبة
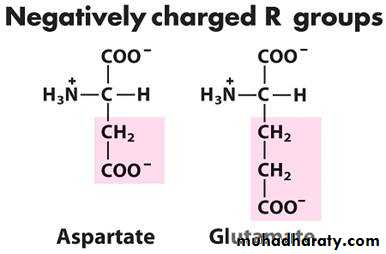
الحوامض الامينية المشحونة بشحنة سالبة24
الحوامض الامينية المشحونة
تقسم الى حوامض مشحونة بشحنة سالبة كما في حامض الكلوتاميك وحامض الاسبارتيك لانها
وعند الرقم الهيدروجيني الفسلجي تكون مشحونة بشحنة سالبة. Amide group تحوي على
اما المشحونة بشحنة موجبة فهي الارجنين والهستدين واللايسين وهي قاعدية التفاعل
والحوامض الامينية المشحونة تكون في الغالب محبة جدا للماء ولذلك توجد في المناطق
الخارجية من البروتين
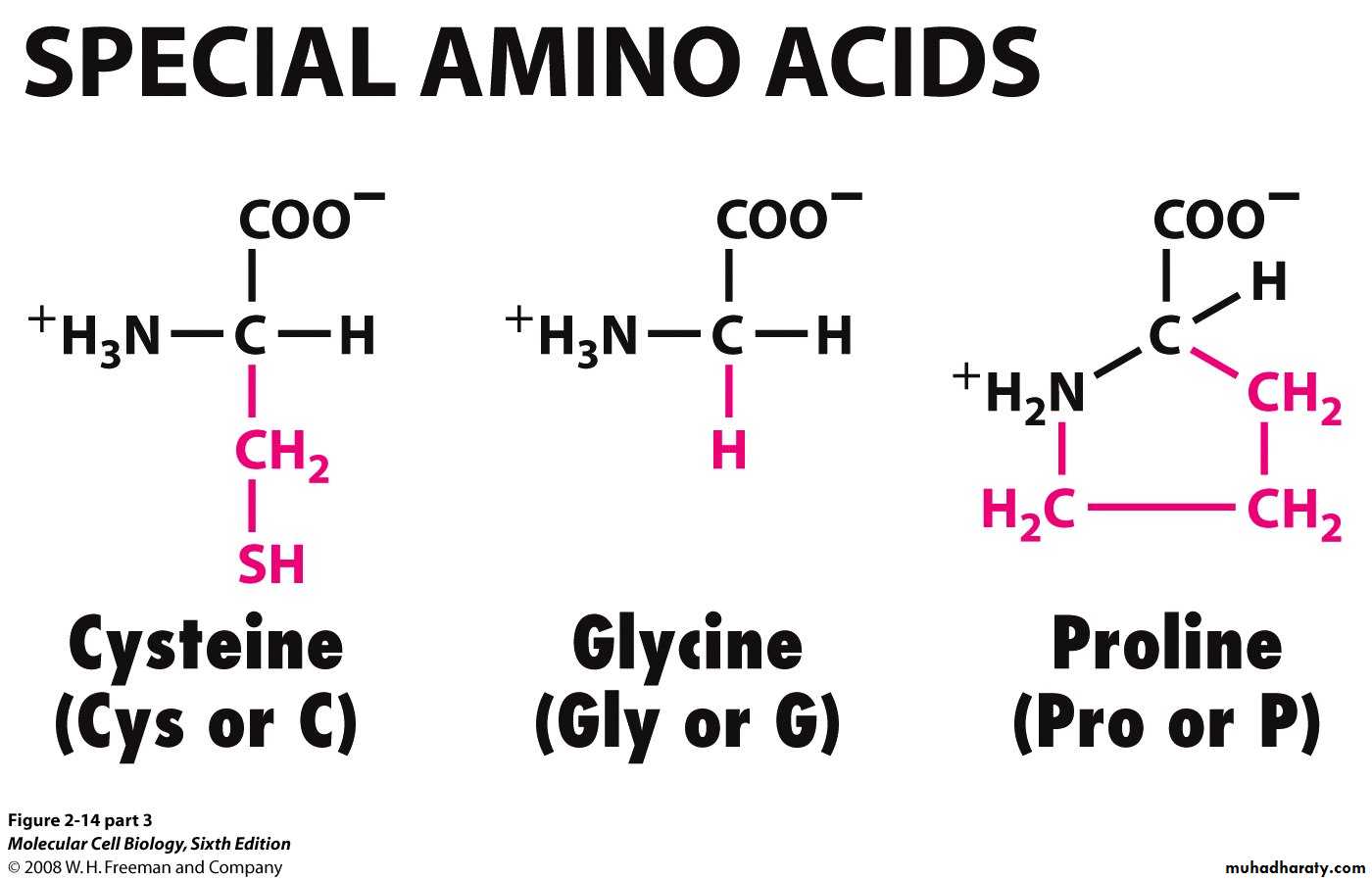
الحوامض الامينية ذات الصفات الخاصة
الملاحظ ان هناك تداخل في تصنيف الحوامض الامينية، فقد يكون أحد الحوامض حاملالصفتين فهو يمكن ان يدخل في مجموعتين. كما ان هناك بعض الحوامض الامينية التي لها
خصوصية مثل الميثايونين والسستئين الحاوي على الكبريت ويكون الأخير أواصر ثنائية
الكبريتيد مع ثمالة مماثلة له في موقع آخر من البروتين ، كما ان الكلايسين يمثل ابسط
الحوامض الامينية كون سلسلته الجانبية مكونة من ذرة هيدروجين فقط، ولذلك يكون أكثر
الحوامض الامينية مرونة ويؤدي الى تكوين تشكيلات تركيبية لا تظهرها الحوامض الأخرى.
وكذا الحال مع البرولين الذي له تركيب خاص فسلسلته الجانبية تكون آصرة مع مجموعة
الأمين الخاصة به مكونا تركيبا حلقيا غير مرن ولذلك لا يمكن ان يوجد في بعض مفاصل
البروتين
25
Imidazole ومن الحوامض الامينية ذات الصفات الخاصة هو الهستيدين الحاوي على حلقة
ويمكن ان يكون متعادلا (بدون شحنة) او مشحونا بشحنة موجبة اعتمادا على البيئة المحيطة،
وهو من الحوامض التي تضفي صفاتها والدفع بها ان تكون في المراكز الفعالة من البروتينات
حيث يقوم بدور تبديل الأواصر وغيرها من الفعاليات.
: ويمكن إجمال تراكيب وبعض صفات الحوامض الامينية بالجدول 8.3
26
الأواصر الببتيدية
وتكون بين ذرات الجزيئة Amide bond الأواصر التي تربط الحوامض الامينية وتسمى ايضا
الرئيسة لتكون سلاسل الببتيدات والبروتينات وتكوين هذه الأواصر سهلا نظرا لطبيعة
خاصة Dipolarized الحوامض الامينية المتأنية وغير المتناظرة اذا انها تكون ثنائية القطب
اذ تكون مجموعة الأمين حاوية على (Zwitterion) عندما تكون في محلول متعادل الحموضة
البروتون والكربوكسيل منزوعة البروتون وهي صفة تعتمد على الرقم الهيدروجيني الذي توجد
فيه الجزيئات وبذلك تمثل مجموعة الكربوكسيل مستقبلا جيدا للأواصر الهيدروجينية ما عدا في
حالة البرولين فان مجموعة الأمين هي التي تكون معطية للهيدروجين ، وعند تكون الآصرة
يرتبط الطرف ألأميني من جزيئة مع الطرف الكربوكسيلي من جزيئة أخرى مع طرح جزيئة
كما موضح في التفاعل الآتي :
27
xxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxxx
مسارات عمليات طوي البروتيناتعمليات طوي البروتينات لا يمكن ان تقتصر على تدخل العوامل الفيزياوية وعلاقتها بالطاقة
التي تشارك بشكل فاعل في الطوي. وفي Chaperones فقط وانما هناك بروتينات الوصيفات
اغلب الأحيان تكون عملية الطوي مرافقة لعملية الترجمة خاصة في البروتينات الكبيرة، وتخليق
البروتينات على الرايبوزوم يبدأ بالطرف ألأميني، والبروتينات الصغيرة تطوى بعد ان يترك
الطرف الكربوكسيلي الرايبوزوم ، اما في البروتينات الكبيرة وخاصة المكونة من عدد من
الدومينات فيبدأ طويها ولا يزال الطرف الكربوكسيلي عالقا بالرايبوزوم .
وعمليات الطوي تكون سريعة وتأخذ مقياس ملي ثانية، فمثلا في البكتريا فان البروتينات تطوى ب 10-50 ملي ثا
عمليات الطوي الخاطئ قد ينشأ من التعرض للحرارة او غيرها من الظروف داخل
الخلية،
28
تراكيب البروتين ومستوياتها
التراكيب البروتينية يمكن ان ترتب هرمياً بأربع مستويات اعتماداً على زيادة التعقيد وهي:التركيب الأولي، التركيب الثانوي، والتركيب الثلاثي المجسم، والتركيب الرباعي.
: موضحة في الشكل 78.3
شكل 78.3 : مستويات تركيب البروتينات
29
وفي العموم تترتب البروتينات على شكلين او صنفين عامة من التراكيب هي: البروتينات
فالكروية هي مطوية وملفوفة . Fibrous proteins والليفية Globular proteins الكروية
بشكل منضغط والتراكيب الليفية تكون على شكل خيوط او أشكال متطاولة، وذلك لوجود مناطق
معينة في الببتيد تحتم الوضع المجسم المحدد الذي يتأثر بتوالي الحوامض الامينية في التركيب
الأولي
-- -
.
شكل 79.3 : تركيب حلزون ألفا
30
Primary Structure التركيب الأولي
ويعني به توالي الحوامض الامينية باتجاه واحد، وتربط الحوامض الامينية مع بعضها بالآصرةالببتيدية ذات الطبيعة التساهمية على الرايبوزوم أثناء عملية الترجمة اي أثناء تخليق البروتين،
ويعد مستوى التركيب هذا أبسط المستويات الموجودة في البروتينات
31
secondary Structure التركيب الثانوي
للآصرة الببتيدية ويتصف بدرجة عالية Local conformation ويعرف ايضا بالهيئة الموقعية
وصفائح a-helixes من التنظيم والترتيب والتكرار. ويتمثل التركيب الثانوي بحلزونات ألفا
ثم تلاها تحديد تراكيب b-sheets بيتا
لتعمل في إعطاء البروتين التركيب W-loop وعروة اوميكا b-turns أخرى مثل لفات بيتا
النهائي.
حلزونات ألفا
تركيب ينتج من التواء السلسلة الببتيدية المفردة، والتوزيع الأساسي للحلزونات يشبه المفك
Cork screw الميكانيكي
تركيبه شبه عصوي يمثل الجزء الداخلي منه السلسلة الرئيسة او العمود الفقري وتبرز السلاسل
: الجانبية الى الخارج كما موضح في الشكل 79.3
32
ويتم تثبيت الحلزون بالآصرة الهيدروجينية بين مجموعة الأمين ومجموعة الكربوكسيل للسلسلة
الرئيسة وتترتب الأواصر الهيدروجينية بحيث تكون السلاسل الجانبية الى الخارج وبصورة
: عمودية على محور التوالي كما في الشكل 80.3
وتكون كل لفة في الحلزون مكونة من 3.6 حامض أميني والمسافة بين اللفات 5.4 انكستروم.
شكل 83.3 : انتظام الأشرطة المكونة لصفائح بيتا
33b-sheets صفائح بيتا
وهي سلاسل ببتيدية متعددة ممتدة وغير b-strands تراكيب ممتدة مكونة من أشرطة بيتا
ملتوية كما في حلزونات ألفا ولذا يكون البعد بين الحوامض الامينية على طول الشريط حوالي
3.5 انكستروم مقارنة ب 1.5 انكستروم في حالة الحلزونات وتكون السلاسل الجانبية موزعة
الى أعلى وأسفل الشريط
وتميل الأشرطة الى إظهار نمط متبادل من المناطق المحبة للماء والأخرى الكارهة له، وعلى
العموم فان الأشرطة المطمورة تكون كارهة للماء وتتكون الصفائح من ارتباط شريطين او أكثر
من أشرطة بيتا بواسطة الأواصر الهيدروجينية، فالأشرطة في صفائح بيتا اما ان تكون متوازية
: او متعاكسة كما في الشكل 83.3
34
شكل 84.3 : المظهر العام للأشرطة في صفائح بيتا
شكل 85.3 : ارتباط صفائح بيتا مع حلزونات ألفا35
التراكيب الأخرى
هناك تراكيب موضعية أخرى لا تعود الى حلزونات ألفا او صفائح بيتا تساهم في تحديدالتركيب الثانوي للبروتينات،
كما في تركيب البروتينات الكروية الذي يحتاج الى عكس الاتجاهات وهذه الانعكاسات تتصف بالعموم
وكذلك عروات اوميكا وانحناءات دبوس الشعر b-turns بكونها استدارات حادة ومنها لفات بيتا
واللفات والعروات تقع في الغالب على سطوح
البروتينات لان ثمالاتها تميل ان تكون من النوع المشحون وثمالات قطبية لذلك فهي تشارك في
التداخل بين البروتينات او تداخل البروتينات مع غيرها من الجزيئات وتنشأ من تكوين الأواصر
الهيدروجينية بين المواقع المختلفة وتكون مهمة من الناحية الوظيفية لانها توجد في العادة في
المواقع الفعالة من البروتين.
: 86.3
شكل 86.3 : اللفات الملتوية
شكل 87.3 : اختلاف قطع من تواليات معينة في إعطاء تراكيب ثانوية مختلفة في بروتينات
مختلفة
36
Supersecondary structure التراكيب الثانوية ألفائقة
تراكيب تتكون من تداخل مكونات التركيب الثانوي بعد ارتباطها بتداخلات مثل الأواصر
الهيدروجينية واستعمال اللفات او الاستدارات او العروات لتكون شكل صلد او منضغط
وهي تراكيب خاصة بحلزونات ألفا Coiled coils ومنها اللفات الملتوية .Compact shape
التي تتصف بتجمع اثنين او أكثر من الحلزونات الملتفة على بعضها كما موضح في الشكل
والحلزونات يكون الطابع الغالب عليها هو كراهية الماء.
وقد تظهر في هذا المستوى تراكيب ثانوية واضحة مرتبة بشكل دومينات ذات وظائف خاصة
وقد تشارك التراكيب الثانوية ألفائقة في التنظيم .helix-turn-helix او قطيفات تركيبية مثل
كما في البكتريا التي تنظم عمليات الانتساخ ببعض من هذه البروتينات ، وكذلك الحال مع
helix-turn-) الذي تركيبه Leucine zipper الأحياء الحقيقية النواة فمثلا زمام الليوسين
او أصبع الزنك تساهم في عمليات تنظيم الانتساخ . (helix
37
Tertiary structure التركيب الثلاثي
التركيب الثلاثي المجسم يمثل مستوى أعقد من المستوى الذي سبقه ويدرس عادة باستعمالالأشعة السينية ومطياف الرنين النووي المغناطيسي. وفي هذا التركيب يصبح هناك توزيعا«
مكانيا" اي تركيب مجسم للتراكيب الثانوية اي ترتب العلاقة المكانية بالنسبة للتراكيب الثانوية
الواحدة مع الأخرى.
وبصورة عامة تكون البروتينات الفعالة هي التي لها تركيب ثلاثي مجسم والذي يتحدد بتوالي
الحوامض الامينية في التركيب الأولي . والشكل التالي ( 88.3 ) يوضح
التركيب المجسم لأحد البروتينات (الأنسولين
):
شكل 88.3 : التركيب المجسم (الثلاثي) لبروتين الأنسولين
.
38
يتم تثبيت التراكيب الثلاثية بواسطة الأواصر الهيدروجينية والتداخلات الأيونية والجسور
الكبريتيدية الثنائية وتداخلات القوى الكهربائية المستقرة وقوى فاندرفال. ولعل أهم هذه القوى
هي الجسور الكبريتيدية ومن
الجدير بالذكر فان هذه الجسور نادرة في البروتينات السايتوبلازمية نظرا لكونها بيئة مختزلة
جدا كما ذكر سابقا في وصف الأنظمة الحيوية
39
التركيب الرباعي Quaternary Structure
يمثل أعقد مستويات التراكيب البروتينية . وهو التركيب الناتج من تداخل عدد من ببتيداتمتعددة في معقد بروتيني تتداخل فيما بينها بتداخلات لا تساهمية وكل ببتيد متعدد يمثل وحدة
والكثير من البروتينات ذات التركيب الرباعي تحوي على اثنين او أكثر Monomer ثانوية او
من الببتيدات المتعددة (الوحدات الثانوية) ترتبط مع بعضها بقوى لا تساهمية تشبه تلك التي
تثبت التراكيب الثلاثية ومنها الجسور الكبريتيدية الثنائية.
.
40
ولذلك فهو يمثل التوزيع المكاني للوحدات الثانوية المتداخلة مع بعضها وأبسط أنواع التراكيب
الذي يتكون من وحدتين قد تكون متشابهة كما في البروتينات Dimer الرباعية هو المزدوج
المكون من Trimer ويليه في التعقيد المكوثر الثلاثي ،DNA التي لها القابلية للارتباط ب
وقد تكون الوحدات الثانوية متشابهة .Multimer ثلاث وحدات ثانوية . والمجمعات الأكبر هي
وتسري التسمية على بقية أعداد الوحدات، ويطلق Heterodimer او مختلفة Homodimer
على الوحدات حتى وان كانت بحدود أربع او خمس وحدات. ومن Oligo- مصطلح قليل التعدد
الأمثلة على البروتينات الفعالة وهي في حالة التركيب الرباعي هو الهيموغلوبين
الذي يحمل الأوكسجين في الدم فهو مكون من وحدتي ألفا ووحدتي بيتا مرتبة Haemoglobin
Hetero-oligomer ولذلك فهو يمثل أحد البروتينات قليلة التعدد المتباينة a2b بشكل رباعي 2
ويكون ترتيب الوحدات في الأغلب له علاقة بمحاور التناظر
41
بعض التراكيب الخاصة في البروتينات
القطيفاتMotifs
تحوي البروتينات على بعض التواليات الخاصة تسمى القطيفات التي يمكن ان تتكرر في توالي
البروتين عدة مرات وتكون في بعض الأحيان بمثابة طمغة مميزة للبروتينات ذات العلاقة
التي توجد في سموم EPIYA وعادة تكون مكونة من توالي حوامض أمينية محددة كما في
او تكون بشكل قطيفات تركيبية مكونة من عناصر . Helicobacter pylori للبكتريا CagA
Inteins الانتينات
من التواليات الأخرى هي الانتينات وتمثل قطع من البروتين تستطيع استئصال نفسها وإعادة
بآصرة ببتيدية ، ولذلك يطلق عليها Exteins لحم او وصل الأجزاء البقية التي يطلق عليها
بعض الأحيان انترونات البروتين.
.mRNA ويعتقد ان الانتينات تتوسط في وصل بعض البروتينات بعد ترجمتها من
42
-
Signal peptide ببتيدات الإشارة
ببتيدات تقع في الطرف ألأميني للبروتين تساعد في1- توجيه البروتينات الى المسار الإفرازي.
وتخلق هذه على الشبكة الاندوبلازمية الخشنة في الخلايا حقيقية النواة. وهناك البعض منها
، (Mitochondria targeting peptide) mtTP 2-يوجه البروتينات الى المايتوكوندريا مثل
في النباتات بإدخال البروتينات (Chloroplast targeting peptide ) cpTP في حين 3- يقوم ببتايد اليخضور
الى داخل البلاستيدات الخضر. 4-وفي الخلايا بدائية النواة فهي تساعد في تصدير البروتينات الى
الخارج عبر الأغشية الخلوية .
.
43
Domains الدومينات
هذه الأجزاء التركيبية هي جزء من البروتين الكلي الذي يثبت نفسه ويطوى بشكل مستقل عنباقي توالي البروتين، ويمكن ان يظهر المتشابه منها في عدد من البروتينات، ويمكن ان تسمى
بشكل منفصل وهي تنفرد بالوظائف الحيوية للبروتينات التي تنتمي اليها مثل دومين ربط
ويمكن ان تقايض او تنقل من بروتين الى آخر
لأداء وظائف جديدة ، وهي الآلية التي تستعمل في العديد من Chimeras لتكوين الكايمرات
البروتينات
44
مواقع البروتينات
تؤدي البروتينات وظائفها في المكان المخصص لها بعد التخليق وإجراء التحويرات اللازمةعليها، فالبعض يبقى في السايتوبلازم او السائل الخلوي والآخر يقتنص في التراكيب الغشائية
وأخرى تفرز الى الوسط المحيط . كما ان بعض البروتينات تحتاجها الخلايا طوال حياتها في
حين هناك بروتينات تحتاجها الخلايا تحت ظروف محددة، كما تختلف الكميات اللازمة في كل
حالة.
45
ومكان وجود البروتينات يكون مقنناً فمثلا المستضدات التي تحفز الجهاز المناعي تكون موجودة
في حين ،Vaccines على سطوح الخلايا مثل البكتريا وتكون مرشحة لإنتاج اللقاحات الوقائية
تكون للبروتينات المطمورة في الأغشية دوراً كبيراً في التأقلم البيئي مثل مكونات الأغشية
اما البروتينات المفروزة فهي التي .Two component systems المكونة من الأنظمة الثنائية
تترك الخلايا بشكل كامل وتكون عادة سامة للخلايا المنتجة لها، او تستعمل لقتل الخلايا
المضيفة فتقوم الخلايا البكترية مثل بحقن المضيف بها مما يؤدي في بعض الحالات الى قتل
المضيف بسرعة . ولكل من البروتينات توالي خاص به يؤهله ان يوجد في المكان الصحيح
فمثلا البروتينات الغشائية تسود فيها الثمالات الكارهة للماء، في حين تحوي البروتينات
او ببتيد الإشارة Signal sequence المفروزة على إشارات خاصة تسمى توالي الإشارة
التي توجد عند الطرف الاميني للبروتين، وإشارات الإفراز هذه هي التي تسحب البروتين الى
الخارج بنظام الإفراز
46
تصنيف البروتينات
تصنف البروتينات لغرض تسهيل دراستها وهو من المزايا المهمة كتوجه عام للإعلان عنالجينومات، وتعتمد عملية التصنيف على عدد من المؤشرات مثل:
التركيب ·
الفعالية ·
الدور الأيضي ·
47
وهناك أكثر من
طريقة لعلمية تصنيف البروتينات منها:
والتي تصنف الى عوائل، Superfamily التصنيف الهرمي لعوائل البروتينات الكبيرة ·
وفي هذه الحالة يعتمد التصنيف على تشابه التواليات ليعطي عناقيد شاملة وغير متداخلة
بشكل ترتيب هرمي الذي يعكس علاقاتها التطورية، والطريقة تسمح بضم البروتينات
الجديدة التي تظهر تشابها" كاملا" للتواليات وبالتالي يشمل التشابه الأطر العامة
للدومينات المشتركة من حيث التركيب او العدد.
بعض العوائل تصنف على أساس التشابه في الدومينات والقطيفات. ·
وفي جميع الأحوال فان علاقة البروتين مع العائلة هي ليست علاقة واحد الى واحد فهي يمكن
ان تكون واحد الى عدة أي ان العائلة يمكن ان تشمل العديد من البروتينات. وتصنف
البروتينات الى عوائل كبيرة ثم عوائل يساعد في تحديد وظائف البروتينات غير معروفة
الوظيفة او ما تسمى بروتينات افتراضية
48
بروتينات خاصة
هناك بعض البروتينات حازت على الاهتمام اما لبساطة تركيبها واستعمالها كموديلات دراسية،البروتينات الخارجية: وتمتاز باحتوائها على الجسور الكبريتيدية الثنائية المكونة من السستئين ·
والتي تكاد تخلو منها البروتينات داخل الخلايا.
البروتينات المعقدة: وتمثل البروتينات التي ترتبط مع جزيئات من هويات أخرى مثل ·
الكربوهيدرات ، وتركيبها هذا ينتج بعد عمليات الترجمة ضمن تحويرات بعد الترجمة ، وتؤثر
في التخصص الوظيفي للبروتينات الناتجة، ويطلق على البروتينات المرتبطة بالكربوهيدرات
Glycoproteins مصطلح البروتينات السكرية
.
49
البروتينات السكرية مثل تلك الموجودة على سطوح كريات الدم الحمر والاختلاف بينها يكون
بالجزء السكري وهي التي تحدد مجاميع الدم التي تزيد عن 100 مجموعة تعود الاختلافات فيها
والتي تخصص بفعالية بعض الجينات المسئولة عن ادماج O, B, A الى الجزء السكري وأهمها
السكر وكذلك في حالة البروتينات السكرية المفروزة .
وتتداخل البروتينات مع الدهون بواسطة تداخلات لا تساهمية لتكون البروتينات الدهنيةlipoprotein
والتي تتعلق وظيفتها بخزن ونقل الدهون والكولسترول في الجسم
50
البروتينات الكروية: تراكيب بروتينية تكون بشكل كريات منضغطة وتكون ثمالاتها الكارهة ·
للماء الى الداخل والمحبة للماء والقطبية الى الخارج لذلك تكون ذائبة ومحاطة بجزيئات الماء،
والهرمونات Cytokines ومحركات الخلايا Myoglobins ومن الأمثلة الشائعة الإنزيمات و
البروتينية.
Ferritin البروتينات المكونة من حلزونات ألفا: من أفضل الأمثلة عليها بروتين الفرتين ·
الخازن للحديد ، و 75 % من البروتين مكون من حلزونات ألفا، والحلزونات فيه يمكن ان
تلتف مع بعضها مكونة تركيب ثابت يصل طوله الى 1000 انكستروم.
وضفائر حلزونات ألفا توجد في بروتينات العضلات والبروتينات الليفية وكرياتين الشعر
والقرون والأظافر، حيث يعمل حبل الحلزونات في مجال الوظائف الميكانيكية بعد ان يكون
حزم من الألياف القوية. وكذلك الحال مع الهيكل الخلوي الذي يمثل دعائم ومنصات الخلية
الداخلية التي تكون غنية بمثل هذه الخيوط.
) .
51
البروتينات الغشائية: تختلف عن باقي البروتينات بكونها توجد في محيط دهني ولغرض ان ·
تثبت في مكانها لابد من ان تتوزع ثمالاتها الكارهة للماء الى الخارج والمحبة للماء الى الداخل. وفي معظم البروتينات التي تستعرض الأغشية تكون تراكيبها الخارجية من حلزونات
ألفا (كما مر ذكره
52
Transcription factors عوامل الانتساخ
بروتينات خلوية تعد الأساس في عمليات التنظيم وذلك بارتباطها الى مواقع محددة علىأي ان mRNA وإنتاج DNA وبالتالي تحدد حركة او انتساخ المعلومات الوراثية من ،DNA
وقد تؤدي المهامة لوحدها او بمساعدة مجموعة أخرى من ، DNA لها القابلية للارتباط ب
او إيقافه mRNA البروتينات وذلك بتكوين المعقدات ويمكن ان تؤدي الى تنشيط تخليق
الى المنطقة المطلوبة. ونظرا لكونها من أساسيات التنظيم لذلك RNAP بواسطة توجيه الإنزيم
يتوقع ان توجد في كل الكائنات الحية. وتختلف أعدادها في الكائنات ولكن الصفة العامة ان
عددها يزداد بازدياد حجم الجينوم ، وفي الجينومات الكبيرة يوجد أكثر من عامل انتساخ \
DNA جين. وفي الإنسان يوجد حوالي 2600 بروتين تحوي على دومينات للارتباط ب
واغلبها من عوامل الانتساخ ، ولذلك فهي تشكل اكبر عائلة بروتينية في الإنسان اذ ان حوالي
%10 من الجينوم البشري يشفر لهذه العوامل.
شكل 89.3 :التركيب العام لعوامل الانتساخ
53
التركيب العام لعوامل الانتساخ
تمتاز معظم عوامل الانتساخ بتركيب أساسي الذي يظهر في الشكل التالي ( 89.3 ) الذي يوضحتركيب البروتين وطرفيه الاميني والكربوكسيلياي
ويحوي البروتين على ثلاث مناطق رئيسة هي:
DNA الدومين الذي يرتبط الى DNA-binding domain = DBD
الدومين الذي يتحسس الإشارات Signal- sensing domain = SSD
الدومين المنشط Transactivation = TAD
54
تصنيف عوامل الانتساخ
هناك أكثر من وسيلة لتصنيف عوامل الانتساخ منها التصنيف على أساس الآلية التي تعمل بها،او الوظيفة التنظيمية او تشابه التوالي.
اولا : تصنيف عوامل الانتساخ بالاعتماد على الآلية منها ما يشترك في تكوين معقدات قبل
وهذه تكون شائعة .TFIIH ، TFIIF ، TFIIE ، TFIID ، TFIIB ، TFIIA البدء مثل
وتتداخل مع منطقة الممهد المحيطة بموقع بدء الانتساخ لمعظم الجينات
وكذلك تشمل عوامل الانتساخ اليسارية وتتمثل بالبروتينات التي ترتبط الى يسار موقع البدء ،
. Basic-Zipper منها أصبع الزنك و
ثانيا : التصنيف الثاني المعتمد على الوظيفة فتقسم عوامل الانتساخ الى:
55
-- -
وتوجد في كل الخلايا وعلى طول Constitutively active TFs عوامل الانتساخ الدائمة ·الوقت.
وهذه تحتاج الى تنشيط وتكثر تحت Conditionally active TFs العوامل الاشتراطية ·
ظروف خاصة.
وتكون هذه في خلايا خاصة Developmental active TFs العوامل الخاصة بالتطور ·
والتعبير عنها يكون تحت سيطرة صارمة وعند بدء التعبير عنها وإنتاجها سوف لا تحتاج الى
تنشيط إضافي.
العوامل المعتمدة على الإشارات وهذه تحتاج الى إشارات خارجية للتنشيط ومنها المستلمات ·
Endocrine او داخلية Paracrine-dependent النووية التي تحتاج الى افرازات خارجية
Autocrine للتنشيط. والبعض الآخر غير النووي يحتاج الى افرازات من داخل الخلية
وغيرها. P مثل 53 dependent
العوامل المقيمة في النواة وهذه تقيم في النواة بغض النظر عن حالتها من التنشيط. ·
وتوجد في السايتوبلازم Latent cytoplasmic factors العوامل السايتوبلازمية الكامنة ·
بصورة غير نشطة وعند تنشيطها تنتقل الى النواة.
.
56
ثالثا : التصنيف الثالث للعوامل يعتمد على تركيبها وتشابه تواليات الحوامض الامينية فيها
وهذه تقسم الى العديد من ،DNA وبالتالي تشابه التركيب الثلاثي للدومينات التي ترتبط الى
الأصناف والأخيرة تقسم الى عوائل كبيرة والتي تقسم بدورها الى عوائل متعددة
57
وظائف عوامل الانتساخ
فارتباطها وعدم ارتباطها يؤثر في ، DNA عوامل الانتساخ يمكن ان تقرأ وتفسر البصمة فيالوظائف الخلوية . وتشارك عوامل الانتساخ في الكائنات متعددة الخلايا في عمليات التطور
والاستجابة للمؤثرات الخارجية. والعوامل الفعالة في هذا المجال تعمل على فتح وغلق الجينات
المطلوبة والتي بالتالي تؤثر في التغيرات الحاصلة في المظهر الخارجي للخلايا، او الفعاليات
التي تحتاجها الخلايا لتقرير مصيرها النهائي الذي يحصل عند تخصص الخلايا و
الجنس.
وتشارك في مهام أخرى مثل حالة التواصل بين الخلايا. فالخلايا تتواصل
بإطلاق جزئيات تعمل على بدء سلسلة من التفاعلات بعد استلامها للإشارات ، ويكون التنفيذ
النهائي لسلسلة نقل الإشارات من مهام عوامل الانتساخ التي تحفز او تثبط العمليات لل
وتشترك العوامل في : .DNA
58
تنظيم الفعاليات في حقيقيات النواة بواسطة عوامل الانتساخ
General في هذه المجموعة من الخلايا هناك مجموعة من عوامل الانتساخ العامة
الضرورية لحدوث الانتساخ، وعدد منها لا يرتبط الى (GTFs) transcription factors
مباشرة. RNA Polmerase وانما يكون أحد مكونات المعقدات الأولية للبدء التي تتداخل مع DNA
وإضافة الى عوامل الانتساخ العامة هناك عوامل أخرى تنظم الجينات بشكل تفاضلي وذلك
المجاورة للجينات المراد تنظيمها. والأخيرة تكون مهمة DNA بالارتباط الى المشجعات من
لتحديد التعبير الذي يحصل في خلايا محددة وفي الوقت المناسب والكميات المناسبة اعتمادا
على التغير في حاجة الكائن لها
59
الاستجابة للظروف الخارجية ·
تعمل عوامل الانتساخ في الاستجابة للظروف الخارجية وتنظم التعبير الجيني على ضوء
الإشارات القادمة ومنها تنظيم الجينات المسئولة عن تخليق بروتينات الصدمة الحرارية، حيث
الذي يكون مهما لتنشيط ( HSF ) Heat shock factor يقوم عامل الصدمة الحرارية
Hypoxia-inducible الجينات اللازمة عند ارتفاع درجات الحرارة. وكذلك العامل الآخر
الذي ينشط الجينات الضرورية لعيش الخلايا في بيئات قليلة الأوكسجين، (HIF) factor
الذي ( SREBP ) Sterol regulatory element binging protein ومثال آخر العاملي بروتين ربط عناصر تنظيم الدهون
يساعد في الحفاظ على مستويات الدهون في الخلايا.
:
60
السيطرة على دورة الخلية ·
والجينات الكابحة Oncogenes هناك بعض عوامل الانتساخ الخاصة بجينات السرطانتساعد في تنظيم دورة الخلية اذ تحدد حجم الخلية Tumor suppressor genes للأورام
c myc وغيرها من المؤشرات لتحديد متى يجب ان تنقسم الخلية الى خليتين بنوية ، ومنها
والذي له دور مهم في نمو الخلايا والاستماتة.
تنظيم الخلايا لعوامل الانتساخ
من المعروف ان الخلايا تحتاج الى مستويات مختلفة من التنظيم وعوامل الانتساخ (التي لها
وظيفة تنظيمية) وعوامل الانتساخ تحتاج الى عمليات تنظيم هي الاخرى وهذه تتم بعوامل
انتساخ أخرى. وتنظيم عوامل الانتساخ يمكن ان يتم بعدة آليات
61
آليات تنظيم عوامل الانتساخ
تخليق عوامل الانتساخ كغيرها من البروتينات التي تنتسخ من جينات على الكروموسومات ·
الذي يترجم الى بروتينات عوامل الانتساخ وأي خطوة من هذه الخطوات تكون RNA الى
عرضة للتنظيم الذي يؤثر في إنتاجها وفعالياتها.
بعض الأحيان تنظم عوامل الانتساخ ذاتيا مثل استعمال عمليات التنظيم الرجعي ·
السلبي فيعمل عامل الانتساخ كابحا لنفسه ويقل إنتاجه بحيث يبقى ضمن الحدود المطلوبة.
62
عوامل الانتساخ كغيرها من البروتينات (في الخلايا حقيقية النواة) تنتسخ في النواة ثم تترجم ·
في السايتوبلازم. وبعض عوامل الانتساخ الموجودة في النواة لها إشارات خاصة تبقيها في
النواة، وهذه تعد مهمة في عملية تنظيم عوامل الانتساخ لانها تؤدي وظيفتها في النواة، لذلك
وجب نقلها من السايتوبلازم بعد تخليقها على الرايبوزومات وهذا يحتاج الى ربطها بربائط
خاصة ثم يتم نقلها الى النواة. ولذلك فان هذه العملية تساعد في التنظيم بمساعدة الدومين
وهذا Ligands باستعمال طرق مختلفة، ومنها الارتباط الى الربائط SSD الحساس للإشارة
الارتباط ليس هو الوحيد الذي يؤدي الى انتقال وحركة عوامل الانتساخ في الخلية. والارتباط
او DNA الى الربيطة يمكن ان يؤثر في عامل الانتساخ وفيما اذا سيكون قادرا للارتباط الى
عوامل مساعدة أخرى.
. DNA الفسفرة لبعض عوامل الانتساخ تكون لازمة لتصبح قادرة على الارتباط الى ·
تتداخل عوامل الانتساخ مع بعضها لتكوين المزدوجات او ترتبط الى بروتينات منظمة ·
مرافقة وبذا تصبح قادرة على أداء وظائفها .
63
الأهمية السريرية لعوامل الانتساخ
لعوامل الانتساخ أهمية سريرية كبيرة وذلك لان:
الطفرات التي تحصل فيها ترتبط بعدد من الأمراض. ·
يمكن ان تكون العوامل أهدافا دوائية. ·
ومن الأمراض المتعلقة بعوامل الانتساخ هو السرطان بشكل رئيس وذلك لتأثيرها في جينات
السرطان او الجينات الكابحة للأورام.
211
شكل 90.3 : تداخل عوامل الانتساخ مع البروتينات الأخرى للقيام بعملية الانتساخ
64
تحويرات بعد الترجمة وأداء البروتينات للوظائف
يبدأ في بعض الأحيان أداء الوظيفة لبعض البروتينات مباشرة بعد تخليقها ولكن في معظمالأحيان تحتاج البروتينات الى عدد من التحويرات التي يطلق عليها تحويرات بعد الترجمة التي
تكون أساسية لأداء الوظائف أي ان البروتين المخلق تواً يكون طليعة لبروتينات أخرى ومن
هذه التحويرات:
طوي البروتين ·
Cofactors إضافة العوامل المرافقة ·
إضافة السكريات ·
إضافة الدهون
وبعض البروتينات تحتاج الى دمج وحدات بروتينية مع بعضها (كما في حالة إنتاج التركيب
الرباعي) لتكون معقدات فعالة. ومن التحويرات الأخرى تكوين الجسور الكبريتيدية الثنائية
والفسفرة وغيرها
65
Epigenetics الوراثة اللاجينية
والهستونات التي تضفي DNA حقل دراسة يتناول التغييرات الكيماوية لكل من المواد الوراثيةالتي تؤثر في الأنماط المظهرية دون المساس بتواليات RNA مرونة على الجينومات ومشاركة
فالجينات يعبر عنها بعد اختيارها وارتباط عوامل الانتساخ بها وفقاً لما تمليه الظروف .DNA
الخارجية التي تحرك الشبكات الخلوية للاستجابة واستعمال تواليات معينة من الجينوم. وارتباط
عوامل الانتساخ الى توالي معين يؤدي الى سلسلة من الأحداث لا تتعلق بتسلسل القواعد
النتروجينية ولكن يتعلق بحاجة الخلية لانتاج بروتين محدد ومهم لها. واي خلل في هذه السلسلة الاشارية يمكن
إحداث السرطانات والعديد من الأمراض الوراثية
فضلا عن مشاركتها في تطور الجينومات ومن هنا يستنتج (ما هو
معروف سابقا) ان:
النمط المظهري = النمط الجيني + مؤثرات البيئة
وتوالياته لا يعني الكل المطلق وانما تساهم بجزء قد يصل الى .% 50 DNA وهذا يعني ان
والجزء الاخر يكون تحت الوراثة اللاجينية او الخارجية وما
والحقل الذي يهتم بدراسته Epigenome يسمى الجينوم الخارجي
وقد كان لتحديد توالي الجينومات الأثر الكبير في تطور الدراسات في هذا الحقل Epigenomics
66
وتتضح معالم مجال الدراسة هذا من حقيقة انه مثلا كل خلايا الإنسان تحمل بصمة
وراثية واحدة ولكن يلاحظ ان الخلايا العصبية تختلف عن خلايا الجلد او اي نسيج او عضو
آخر.
ويظهر تأثير الوراثة اللاجينية في المراحل المختلفة من الحياة وكذلك الاختلاف في الجنس
والأنماط المظهرية التي تظهر بتأثير ظروف المناخ والغذاء وغيرها. وتشير الدراسات الى ان
في البشر متشابه الى حد % 99.9 وان الاختلافات 0.1 % تعزى الى تغايرات DNA تواليات
المارة الذكر فضلا عن مساهمة الوراثة اللاجينية في ذلك. ومن هنا SNPs النيوكلوتيد المفرد
يلاحظ ان الوراثة اللاجينية تدمج كل اللغات الكيماوية التي يستعملها الجينوم والبيئة للتواصل
وبالتالي فهم التطور ودراسة الحالات الطبيعية من الحالات المرضية
67
Prions البريونات
تمثل أحد عوامل الوراثة الخارجية اللاجينية وهي أحد أشكال البروتينات المسببة للإصابة
بالأمراض. وحقيقتها انها بروتينات مطوية بشكل غير صحيح تؤدي الى تخريب الوظائف
الخلوية وفي الفطريات والخمائر تستورث هذه المكونات دون التأثير في الجينوم . وتأثيرها
يكون من خلال حجزها البروتينات في تجمعات خاصة والبعض منها يزيد من عملية الترجمة
بالمساعدة بعبور شفرات الوقف وبالتالي اما تؤدي الى التخلص من الطفرات التافهة او تزيد من
طول البروتين عن المقنن له.
68
.
Bioinformatics and the internet المعلوماتية الحيوية والانترنيت
يمثل علم المعلوماتية الحيوية مجموعة متوافقة من العلوم والتي تنمو باستمرار وذلك باستخدام
طرق حسابية (عددية) للإجابة على الأسئلة البايولوجية. وتتطلب الإجابة على هذه الأسئلة
الاستفادة من مجموعة البيانات الكبيرة والمعقدة للوصول الى الاستنتاجات البايولوجية.
يلعب حقل المعلوماتية الحيوية مع الاكتشافات الكبيرة للمعلومات الخاصة بالتواليات والتراكيب
الخاصة بها المتاحة للباحثين دورا متزايدا في دراسة أساس المشاكل الطبية الحيوية. والسؤال المهم هنااو
(التحدي) هو كيفية تقديم المساعدة في اكتشاف الجين وغيرها من المعلومات ، والتجارب من
الأنواع الأخرى التي يمكن ان توحي الى احتمالية وجود علاقات غير معروفة مسبقا نسبة الى
تركيب ووظيفة كل من الجين والبروتين.
وللاجابة على ذلك لابد من استخدام كل الجوانب العلمية للعديد من العلوم لتحديد مثل هذه العلاقة غير معروفة مسبقا
ومن احد هذه الجوانب العلمية المهمة المفاهيم الاحصائية.
69
statistical concepts
Normal distribution التوزيع الطبيعي
عندما يتم Continuous random variable يظهر مصطلح المتغير العشوائي المستمر
التعامل مع الكميات التي تقاس بمقاييس مستمرة. على سبيل المثال عند قياس عمر شخص
معين او الوزن ....الخ. في الجانب العملي غالبا ما يتم تدوير القيم الى اقرب عدد صحيح،
بعبارة أخرى يمكن ان يقال ان المتغير (تحت الدراسة) متغير مستمر في حالة كون القيم التي
يتعامل معها هي قيم مستمرة وتحدد بفترات. في هذه الحالة (الحالة المستمرة) يشار الى
الاحتمالية من خلال مفهوم المساحة.
ان المنحنيات المستمرة عبارة عن أشكال بيانية ناتجة عن دوال والتي يشار اليها بالتوزيعات
المستمرة . يرمز الى التوزيعات الاحتمالية المستمرة بالمساحة تحت المنحني بين قيمتين على
( شالتوزيع كل 11.4 )،b و a سبيل المثال
b و a شكل 11.4 : المنحنى الطبيعي فيه المساحة تحت المنحنى بين
شكل 11.4 : المنحنى الطبيعي فيه المساحة تحت المنحنى بين
Gaussian ان التوزيع الطبيعي او في بعض الأحيان يشار اليه بتوزيع كاوسيان
له أهمية كبيرة وذلك : distribution
-1 له العديد من التطبيقات في الواقع العملي.
70
-2 ان العديد من متوسطات العينات التي لها توزيعات تتبع التوزيع الطبيعي، وذلك من
خلال إجراء التقارب بين هذه التوزيعات والتوزيع الطبيعي.
للمتغير العشوائي المستمر Probability density function(pdf) ان دالة الكثافة الاحتمالية
وكالاتي:، x هي عبارة عن صيغة لإيجاد احتمالية X
ثابت قيمته p هو الانحراف المعياري للتوزيع الطبيعي، و s هو المتوسط و m حيث ان
.
شكل 12.4 : توزيع المنحنى الطبيعي وفيه المساحة مقسمة حسب المقياسين الطبيعي والقياسي
ان الميزة المهمة للتوزيع الطبيعي ان المعادلة الرياضية يمكن تحديدها بصورة تامة اذا عرفت
قيمتي الوسط والانحراف المعياري .
من الجانب العملي، يتم استخراج المساحة تحت المنحني للرسم البياني للتوزيع الطبيعي
لغرض استخدام هذا الجدول يجب التحويل من . (I الجدول ملحق ) z بالاستعانة بجدول قيم
Standard normal distribution الصيغة الطبيعية (التوزيع الطبيعي) الى التوزيع القياسي الطبيعي
وكالاتي:
Z=x- Mu/ s
71
مقاييس النزعة المركزية Measures of central tendency
غالبا ما تكون هنالك ضرورة من تمثيل مجموعة من البيانات بمقاييس متمثلة برقم مفرد واحدوهذا الرقم يتمركز في منتصف البيانات ليعبر عنها. وان أهم هذه المقاييس هي المتوسط
والوسيط والمنوال.
Mean , Median , and Mode المتوسط والوسيط والمنوال
عند وصف البيانات ، يتجه التفكير الى قيمة المتوسط او المعدل او القيمة التي تتوسط البيانات
للتعبير عنها. هنالك ثلاثة أنواع من مقاييس النزعة المركزية. المتوسط ( ) وهو عبارة عن
مجموع جميع القيم المشاهدة للمتغير (تحت الدراسة) مقسوم على عدد المشاهدات
72
Systolic blood pressure مثال 15.4 : الجدول ( 5.4 ) يمثل قيم انقباض ضغط الدم
116 116 115 110 105 104 100 100
128 128 124 122 120 120 120 116
138 134 130 130 130 130 130 130
144 140 140 140 140 140 140 140
170 170 152 150 150 150 150 144
احسب معدل انقباض ضغط الدم ل 40 مريض.
المعدل
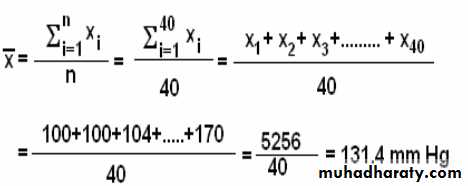
73
ان الشهرة التي يتمتع بها مقياس المتوسط او المعدل كونه مقياس الوسط او المركز لمجموعة
من البيانات ، لم تكن صفة عرضية ، إضافة الى ذلك فهو مقياس بسيط وله العديد من
الخصائص الآتية:
-1 يمكن حسابه لأية مجموعة رقمية من البيانات، وبهذا فانه موجود دائما.
-2 كل مجموعة رقمية من البيانات لها معدل واحد وواحد فقط ، وبهذا فان هذا المقياس
هو وحيد دائما.
-3 يمكن إجراء معالجات حسابية أخرى او إضافية عليه. مثال ذلك يمكن حساب
المعدل العام لمجموعة من المعدلات المختلفة المحسوبة من عدد من المجموعات المختلفة
بعبارة أخرى احتساب معدل المعدلات.
-4 يأخذ في حساباته كل عنصر في البيانات ، وهو بالتالي لا يهمل أية قيمة مهما كانت
صغيرة او كبيرة.
-5 يمكن الوثوق نسبيا في حالة احتساب عدد من المعدلات الخاصة بعدد من العينات
المختلفة المسحوبة من مجتمع واحد بان تكون قيمها (قيم المتوسطات) غير متذبذبة او
مختلفة اختلافا كبيرا.
مثال( 16.4 ): أعمار سبعة طلاب هي 20 , 18 , 19 , , 17, 20 , 19 , 18 , وكان عمر
التدريسي الذي يدرسهم 53 . فان معدل الأشخاص الثمانية هو :
74
Data analysis تحليل البيانات
تحليل البيانات هو التطبيق الحقيقي الذي يحدث في مجالات المعلوماتية الحيوية ولكن قبل البدءلابد من معرفة ان الأنظمة الحيوية أنظمة معقدة جدا وترتبط الجزيئات والتراكيب الخلوية فيما بينها بشبكات
وأخرى شبكات صغيرة Global networks معقدة جدا، قد يقسمها البعض الى شبكات عامة
Minor networks او ثانوية
اي ان الأنظمة الحية او الخلايا لا تكون بمعزل عن المحيط
الخارجي اي ان الخلايا تخضع للقوانين الكيماوية والفيزياوية على حد سواء، وبذا يمكن القول
هي الأعقد على وجه البسيطة، ولكن ما لا يدرك كله لا Bionetworks ان الشبكات الحيوية
يترك جله ومن هنا نشأت محاولات الدراسة لفهم هذه الأنظمة.
المشكلة الأخرى والناتجة عن تعقيد الأنظمة الحيوية هي ان المواد الوراثية تمثل المستوى الاول
، A,C,G,T,U من مستويات الخلية تتمثل بأربعة الى خمسة حروف هي القواعد النتروجينية
في حين ان التطبيقات في المستوى الثاني يكون 20 الى 21 حامض أميني، كما ان القضية
المحورية التي افترضت في بدايات دراسة علم الحياة الجزيئي والمتمثلة بان مكنون الجينوم او
يحول الى مكنون بروتيني. RNA ينتسخ ليكون مكنون النسخ المتمثل ب DNA
كل ذلك عقد فهم المعاومات فظهرت الحاجة لتحليل البيانات لتسهيل فهمها.
75
تطبيق عمليات التحليل
لمعلومات يقوم المستخدم بإعدادها او Execution pipeline معظم التحليلات هي سبل تنفيذية
الحصول عليها من مراجع أكاديمية او مصادر تجارية واختيار طريقة التحليل تؤثر بشكل كبير
في نوعية النتائج التي يتم الحصول عليها. وتتمثل عملية تنفيذ طرق التحليل بشبكات متداخلة
الخطوات، في كل خطوة يتم أداء جزء من المهامة مثل إجراء الحسابات على البيانات المدخلة
وغيرها من العمليات. والشكل التالي( 1.5 ) يوضح بعض الطرق التنفيذية المستعملة:
76
فيلاحظ ان عمليات المعالجة تكون متسلسلة في الجزء الأيسر وفيها ان الخطوة الثانية (مثلا) لا يمكن ان تبدأ إلا بعد اكتمال الخطوة الأولى لتكون معطياتها مجالا لتطبيق الخطوة الثانية
وهكذا الحال مع الخطوة الثالثة ، في حين ان الطريقة الثانية (الجزء الأوسط) يلاحظ ان
العمليات تسير بشكل متزامن في البداية لتعطي معطيات للمرحلة الأخيرة ثم إعطاء النتائج
ويستخدم الحاسوب في تنفيذ كل عملية وهذا التزامن يمكن ان يؤدي الى بطيء الحاسبة والتنفيذ.
اما النوع الثالث فتكون المدخلات فيه متعددة (الجزء الأيمن) وتسير العمليات بشكل متواز وهي
الطريقة الأكثر استعمالا في بعض مجالات المعلوماتية الحيوية كما في البحث عن الموائمة
.BLAST للبروتينات او الجينات في قواعد المعلومات عند استعمال برنامج Matching
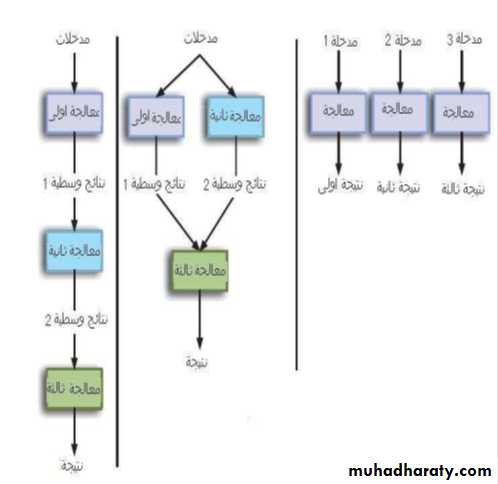
شكل 1.5 : طرق تنفيذ تحليل البيانات
77تحديد تشابه التواليات
نظرا للتعقيدات المذكورة أعلاه ، لابد منتدخل طرق الإحصاء ووضع المصفوفات لغرض ترجيح صحة النتائج والتي تبقي على هامش
ولو قليل من الشك نظرا لتعقيد الأنظمة الحيوية.
Alignment الاصطفاف
أبسط حالاتها. وتعد Pairwise sequence alignment يمثل اصطفاف زوج من التواليات
مقارنة التواليات أحد أهم التحاليل المستخدمة في المعلوماتية الحيوية، فهي الخطوة الأولى في
اتجاه تحليل الهيكل والوظيفة للتواليات الجديدة . وفيها يتم البحث عن رموز ثمالات الحوامض
وتعد عملية اصطفاف . DNA الامينية المتماثلة في التواليات ذات العلاقة او رموز قواعد
، Similarity زوج من التواليات الأساس في البحث في قواعد البيانات عن التشابه للتوالي
.Multiple sequence alignment وكذلك الأساس في عمليات الاصطفاف لعدة تواليات
وتوفر عملية اصطفاف التواليات إمكانية الاستدلال على العلاقة بين التواليين تحت الدراسة.
.
78
فانه من المستبعد للغاية ان Identity فاذا أشتراك التواليان بدرجة معنوية من التشابه او التماثل
يكون التطابق عشوائيا، وهذا يعني ان التواليين يمكن ان يكونا منحدرين من أصل تطوري
ولكن يكون هناك احتمال لعدم التطابق في بعض ، Homologous ( مشترك (التماثل الجذري
المواضع ناتجا عن عمليات استبدال الحوامض الامينية (في حالة البروتينات) وهنا يمكن ترجيح
احتمال انحدار التواليان من أصل مشترك ولكن تباعدا بحيث أصبحت العلاقات الجذرية غير
قابلة للتميز على مستوى التوالي
79
التماثل الجذري والتشابه والتماثل للتواليات
أحد الأسس المهمة في تحليل التواليات، Sequence homology يعد التماثل الجذري للتوالياتفعندما ينحدر تواليان من أصل مشترك فيمكن القول إنهما يمتلكان تماثلا" جذريا" او يشتركان
في جد مشترك، في المقابل يمثل مصطلح تشابه التوالي:هو نسبة اصطفاف وموائمة ثمالات
الحوامض الامينية التي هي متشابهة في الخصائص الفيزوكيمياوية مثل الحجم والشحنة وكرهها
للماء، لذا كان من المهم التمييز بين مصطلح التماثل الجذري والمصطلح المتصل به وهو تشابه
اي ان الثمالات Sequence identity وكذلك تماثل التوالي Sequence similarity التوالي
تكون هي نفسها في التواليين وليس من المجموعة، والحقيقة ان هناك خلط في استعمال هذه
المصطلحات.
.
80
فالتماثل الجذري يستدل به من علاقة الأسلاف المشتركة المأخوذة من مقارنة التواليات ويكون
التواليان بدرجة عالية من التشابه. اما مصطلح التشابه فهو نتيجة مباشرة من المشاهدات الناتجة
من اصطفاف التواليات. ويمكن تحديد كمية تشابه التواليات باستخدام النسب المئوية في حين ان
التماثل الجذري للتواليات هو حالة نوعية، فعلى سبيل المثال يمكن القول ان التواليين يشتركان
بنسبة 40 % من التشابه ومن غير الصحيح القول ان التواليين يشتركان بنسبة 40 % من التماثل
الجذري فهما اما ان يكون هنالك تماثل جذري بين التواليين او لا يكون. وعلى العموم اذا كان
مستوى تشابه التواليان عاليا" بما فيه الكفاية يمكن ان يستنتج منه ان هناك علاقة تطورية
مشتركة، ولو انه في بعض الأحيان تكون العلاقة غير واضحة دائما، ويعتمد الجواب على نوع
التوالي تحت الدراسة وعلى طول التواليات
81
فعلى سبيل المثال لو تم صف تواليين بطول 100 حامض أميني فان التشابه بنسبة 30 % او
أعلى يدل على ان التواليين يمتلكان تشابه متقارب. وعند تمثيل الأهمية الإحصائية تظهر في
الآتي
شكل 2.5 : التمثيل الإحصائي لدرجات التشابه
30 % يؤشر الى ان علاقة التشابه تقع ضمن المنطقة غير المؤكدة - فان مستوى التطابق بين 20
اذ يتداخل فيها التشابه الضئيل Twilight zone والتي يطلق عليها منطقة الغروب او الغبش
مع تداخل التواليات نتيجة للعلاقات العشوائية. اما عندما تكون نسبة التشابه اقل من 20 % فان
نسبة عالية من التواليات تظهر بأنها تواليات لا علاقة بينها، وتظهر ممثلة في الشكل بما يطلق
وهي نتائج لا يمكن الاعتماد عليهاMidnight zone عليه منطقة الظلام او غير مقبولة
ولذلك احتاجت هذه النتائج الى طرق إحصائية أكثر صرامة لتحديد
علاقات التشابه لتحديد المعنوية الإحصائية .
شكل 2.5 : التمثيل الإحصائي لدرجات التشابه
82يتم استخدام التماثل والتشابه بصورة مترادفة في التواليات النيوكلوتيدية ولكن هذه تكون مختلفة
في توالي البروتينات ، اذ يشير تماثل التواليين الى النسبة المئوية لوجود الحوامض الامينية
نفسها في التواليين، في حين يشير التشابه الى النسبة المئوية لاصطفاف الحوامض الامينية من
الزمرة ذاتها اي التي تمتلك خصائص فيزيوكيماوية متشابهة والتي تكون أكثر استعداداً
للاستبدال بعضها مع البعض الآخر دون تأثير كبير.
83
وهناك طريقتان لاحتساب التماثل والتشابه للتواليات تتضمن :
الأولى: استخدام أطوال كلا التواليين.
وبلغة الرموز تستخدم هذه الطريقة الصيغة الآتية:
S= [(Ls × 2) / (La + Lb)] × 100
a , b تمثل النسبة المئوية لتشابه تواليان S حيث ان
عدد الأحماض الامينية المصطفة المتشابه الخصائص Ls
المجموع الكلي لأطوال لكلا التواليين على انفراد . Lb و La
بطريقة مماثلة وكما يأتي: (I%) ويحتسب تشابه التوالي
I = [(Li × 2) / (La + Lb)] × 100هو عدد الأحماض الامينية المتماثلة. Li حيث ان
الثانية: تعتمد على النسب المئوية لاعداد من الثمالات المتماثلة والمتشابهة منسوبة الى المجموع
الكلي للثمالات في التوالي الأقصر.
تعتمد هذه الطريقة في الحساب النسبة المئوية لكل من تماثل وتشابه أعداد الأحماض الامينية
على مجموع الأحماض الامينية للتوالي الأقصر وباستخدام الصيغة:
S= [(Ls × 2) / (La + Lb)] × 100
I(S)% = Li(S) / La %
طول التوالي الأقصر من التواليين تحت الدراسة . La حيث ان
84
مثال : بافتراض التواليين التالية والمراد احتساب :
لهما .(عدد المتشابهات 7) Similarity أ – التشابهلهما .(عدد المتماثلات = 2) Identity ب – التماثل
Seq 1a:CRSCRHAPSRMLAACGAPV
(b) Seq 2b: AAAAVPCTLIMLIRSCGAAGCFTIL
وفق مواصفات الحوامض الامينية التي تم تقسيمها الى عدة مجاميع كما في المخطط الآتي:
أ – يمكن احتساب التشابه من المعادلة الآتية :
S= [(Ls × 2) / (La + Lb)] × 100
S=[(7 × 2 ) / ( 20 + 25)] × 100
S= 31.1%
او باحتسابها وفق المعادلة القياسية وكالاتي :
S = ( Ls / La ) × 100
S = ( 7 / 20 ) × 100
S = 35%
85
-
او باستعمال المعادلة القياسية وكالاتي :
I = ( Li / La ) × 100
= ( 2 /20 ) × 100
= 10%
من المعادلة الآتية : Identity ب – يمكن احتساب التماثل
I = [(Li × 2) / (La + Lb)] × 100
I = [ ( 2 × 2 ) / ( 20 + 25 ) × 100
86
مثال : بافتراض ان التواليين التالية والمراد حساب
لهما علما ان اللاحماض الامينية المتشابهة بالصفا ت= 10 . Similarity أ – التشابهلهما (علما ان المتماثلات 2) . Identity ب – التماثل
(a) Seq 1 : AAAAVPCTLIMLIRSCGAAGCFTIL
(b) Seq 2: CRSCRHAPSRMLAACGAPVALMFPA
وفق المخطط أعلاه لمجاميع الحوامض الامينية يمكن
أ – حساب التشابه وفق المعادلة الآتية
S = [(Ls × 2) / (La + Lb)] × 100
S = [(10 × 2 ) / ( 25 + 25 )] × 100
= 40%
او حسابها وفق المعادلة القياسية الآتية :
S = ( Ls / La ) × 100
= ( 10/25 ) × 100
= 40%
ب – يمكن حساب التماثل من المعادلة
I = [(Li × 2) / (La + Lb)] × 100
= [ ( 2 × 2 ) / ( 25 + 25 ) × 100
= 8%
او حسابها وفق المعادلة القياسية الآتية :
I = ( Li / La ) × 100
= ( 2 /25 ) × 100
= 8%
87
طرق اصطفاف زوج من التواليات
الهدف العام من صف زوج من التواليات هو إيجاد أفضل زوج من التواليات يمتلكان اكبر عدد
ممكن من تشابه الحوامض الامينية وتمثل إحدى التواليات توالي الاستعلام المراد البحث عن
والتي تكون معلومة التركيب والوظيفة. Subject sequence توالي يشبهه في قواعد البيانات
ولذلك يتم صف التواليين وتكون عملية الاصطفاف على نوعين:
وهو الأكثر استعمالا. Global alignment الاصطفاف الشامل ·
. Local alignment الاصطفاف الموضعي
في الحالة الأولى يتم صف التواليين الذي يفترض ان يكونا متساويين في الطول، وينفذ
الاصطفاف من بداية التوالي الى نهايته لإيجاد أفضل تلاؤم ممكن لهما عبر الطول الكلي
للتوالي. وهذه الطريقة أكثر تطبيقا لصف تواليين متقاربين في العلاقة ومتساويان في الطول.
وفي حالة التواليات المتباعدة او غير المتساوية في الطول فان الطريقة غير قادرة على إيجاد
أفضل النتائج لانها لا تستطيع تمييز المناطق الموضعية ذات التشابه العالي بين التواليين،
: وطريقة الصف الشامل موضحة في الشكل 3.5
شكل 3.5 :اصطفاف التوالي الشامل
متساوية الطول
88
ويشير (:) الى تماثل الثمالات في حين(.) يشير الى تشابه الثمالات اي ان الثمالات ليست
نفسها ولكن من زمرة واحدة في الحوامض الامينية.
اما في حالة الاصطفاف الموضعي فلا تحتاج التواليات ان تكون متساوية الطول ، والطريقة
تعمد الى إيجاد المناطق الموضعية التي على مستوى عالي من التماثل دون الأخذ بنظر الاعتبار
اصطفاف بقية المناطق في التوالي، وتستخدم هذه الطريقة لصف التواليات غير متساوية الطول
: والطريقة موضحة في شكل 4.5

شكل 4.5 : اصطفاف التوالي الموضعي
غير متساوية الطول
89
خوارزميات الاصطفاف
خوارزميات الاصطفاف الشامل والموضعي تكون متشابهة في الأساس وتختلف في إستراتيجيةتحديد الامثلية في اصطفاف الحوامض الامينية المتماثلة، وكلا النوعين يمكن ان يعتمد على
واحدة من الطرق الآتية:
Dot matrix method طريقة مصفوفة التنقيط ·
Dynamic programming method طريقة البرمجة الداينميكية ·
Word method طريقة الكلمة ·
-- طريقة مصفوفة التنقيط : وتعد من الطرق المهمة في صف التواليات وتعرف ايضا بطريقة
والطريقة تصويرية تستخدم لغرض المقارنة بين تواليين في .Dot plot map خارطة التنقيط
مصفوفة ذات بعدين، وفيها يكتب التواليان المراد المقارنة بينهما على المحورين السيني
والصادي من المصفوفة. ويتم المسح لكل قاعدة لأحد التواليان مع ما يماثلها من القواعد في
التوالي الآخر، وعند وجود تطابق لأحد القواعد فانه يحدد بنقطة على الخارطة او المصفوفة،
وفيما عدا ذلك يترك الموضع شاغراً، وعند تحديد مناطق التطابق لكلا التواليين تظهر خطوط
من نقاط تصل بينها بشكل خطوط قطرية متجاورة والتي تشير الى اصطفاف التواليات كما
: موضح في الشكل 5.5
شكل 5.5 : طريقة مصفوفة التنقيط للاصطفاف
90
شكل 6.5 : ظهور القطع وعدم التواصل نتيجة لحدوث الطفرات او عمليات الحذف او الإضافة
وتستخدم الطريقة لتحديد التكرار الداخلي للعناصر المكونة للتوالي نفسه وعندها يكون هناك
قطر رئيس يمثل التطابق التام، وعند وجود المكررات مثلا في توالي ما يلاحظ ظهور خطوط
: قصيرة متوازية فوق وتحت الخط او القطر الرئيس موضحة في الشكل 7.5
شكل 7.5 : ظهور المكررات في عمليات الاصطفاف
وطريقة مصفوفة التنقيط للاصطفاف الموضحة في الشكل ( 6.5 ) لمقارنة تواليين ، فتشير
الخطوط على القطر الرئيس الى اصطفاف التواليين أي تماثلهما ، وتشير الخطوط فوق وتحت
القطر الرئيس الى التكرار لكلا التواليين ، وعند ظهور قطع في الخط القطري دل ذلك على
وجود حالات إضافة او حذف، اما الخطوط القطرية المتوازية داخل المصفوفة فتمثل المناطق
: المتكررة من التوالي. كما موضح في الشكل 6.5
91
وعند تمثيل التطابق بالبرامج التصويرية تظهر بشكل مستوي كما موضح في نتائج البرنامج
: ( لصف البروتينات (شكل 8.5 Clustal X
Clustal X شكل 8.5 : نتائج صف تواليات باستعمال برنامج
ولطريقة مصفوفات التنقيط تطبيقات في التعامل مع الجينومات حيث تستعمل في تحديد
المكررات في الكروموسومات التي تظهر بشكل خطوط متوازية بالطول نفسه وكذلك تستعمل
في تحديد التراكيب الثانوية للحوامض النووية وذلك بالكشف عن التواليات المكملة.
92
: Global Sequence alignment
يمكن عمل اصطفاف تواليان بشكل يدوي بدلا عن الكومبيوتر وكما يلي:
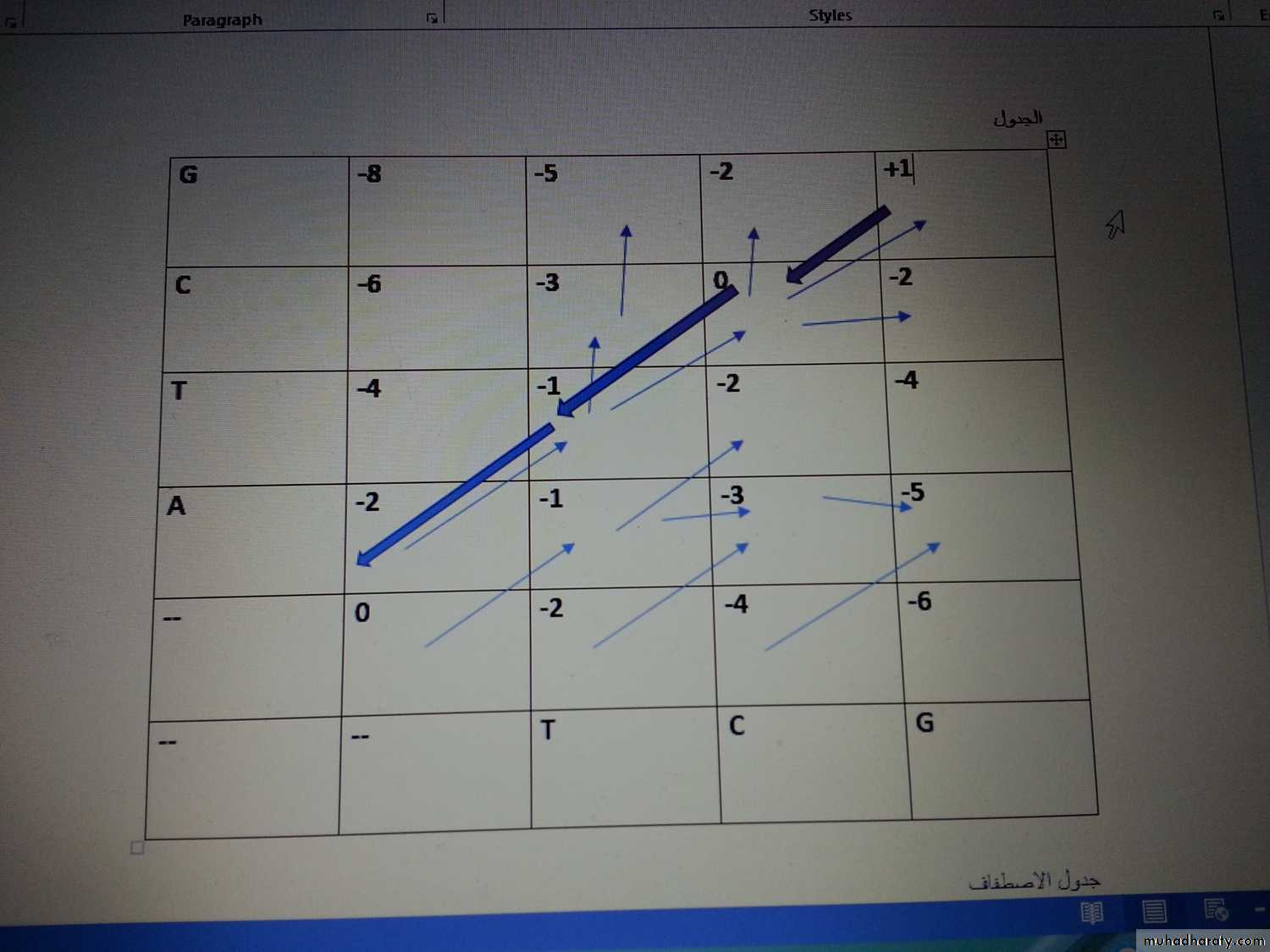
مثال: بين كيف يتم صف التواليين ادناه بطريقة الاصطفاف الشامل وحسب القيم المعطاة ( Match=1 , Mismatch= -1 , Gap= -2 )
التوالي : Seq1=ATCG
Seq2= TCG
خطوات العمل
1-نرسم جدول بعدد حروف التوالي تم نوزع الحروف (التوالي العلوي نضعة عموديا والتوالي السفلي نضعه افقيا.
2-نضع فجوة في البداية (عند بداية اول عمود واول صف) لان ربط متوالتين تحتاج وضع فراغ في البداية والفراغ يعبر عنها في الجدول بقيمة صفر.
3-نضع القيم (scoring)
نضع القيم المعطاة اعلاه بالجدول وكما يلي Match=1 , Mismatch= -1 , Gap= -2 )
93
94
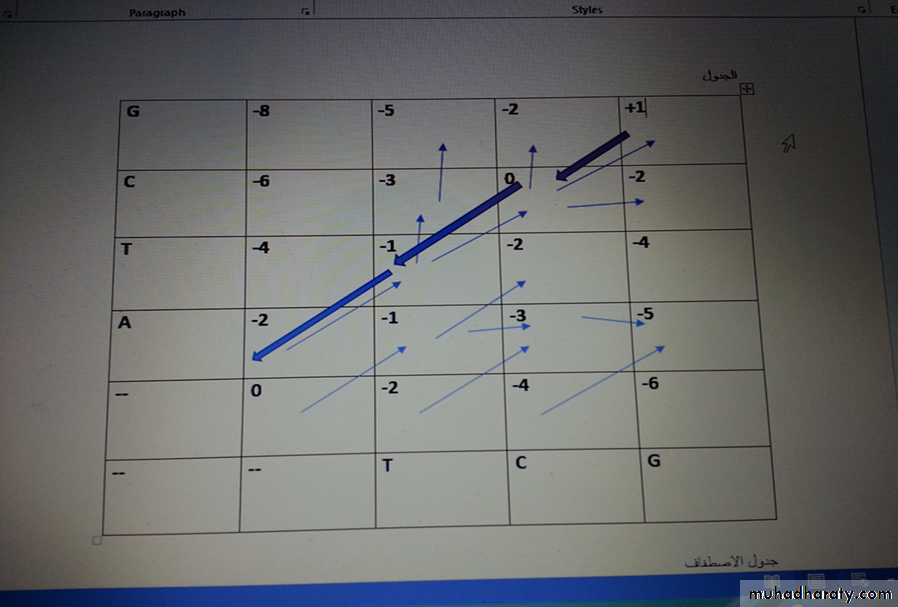
كيفية حساب قيمة كل مربع
ملاحظة مهمة جدا انه عند حساب كل قيمة من الثلاث قيم نظيف قيمة الفجوة العامة لكل رقم ياتي من كل مربع من المربعات الثلاثة المجاورة للمربع الذي نريد ايجاد قيمته ويحسب كما يلي -1-بداية نبدأ بالصف الاول : الذي يبدأ بمربع محاط بثلاث مربعات قيمتها صفر (فا رغة) فقيمته صفر
المربع الذي يليه يحسب كما يلي : على يساره صفر + قيمة الفجوة معلومة -2) اذن قيمته = -2
المربع الذي يليه : القيمة على يساره -2 + قيمة الفجوة -2 = -4
المربع الذي يليه: على يساره -4 + قيمة الفجوة -2 = -6
2- ناتي على العمود الاول : المربع الاول يقابل للحرف A ويقابل فجوة وقيمتها -2 اذن يعطى -2
المربع الاعلى 0المواجه لحرف ال T ومن جهة اخرى يواجه قيمة فجوة -2 , يضاف له قيمة فجوة عامة -2 اذن النتيجة -4
المربع الاعلى (مواجه للحرف C )من اسفله قيمة فجوة -4 + قيمة الفجوة العامة -2 = -6
المربع الاخير : من الاسفل -6 + قيمة الفجوة العامة -2 = - 8
95
3- ناتي على المربع الاخر (المواجه للحرف A بالعمود والمواجه للحرف T من الصف
من اليسار القيمة -2+قيمة الفجوة -2 =-4ومن الاسفل القيمة -2+ قيمة الفجوة -2 =-4ومن المحور القيمة تستخرج من حالة التطابق (+1) اوغير التطابق (-1) او الفجوة (-2) وبما ان A غير مطابق ل Tاذن ياخذ قيمة -1, اذن اصبح لدينا ثلاث قيم ناخذ اعلى قيمة ونثبتها على المربع وهي قيمة غير مطابق -1 ونثبت سهم محوري يظهر مكانها (اي مصدرها)
4- المربع الذي يليه هو المواجه للحرف A بالعمود والحرف C بالصف وقيمته كما يلي وبنفس الطريقة:
من اليسار -1 + قيمة الفجوة العامة -2 = -3
من الاسفل -4 + قيمة الفجوة العامة -2 =-6
من المحوري (عدم تطابق اي مع سي ) -1 + قيمة الفجوة العامة -2 = -3 , اذن نثبت اقل قيمة وهي -3 ونثبتها بالمربع ونثبت مكان صدورها 0سهمين من المحور ومن اليسار). وهكذا نفعل مع بقية المربعات جميعها
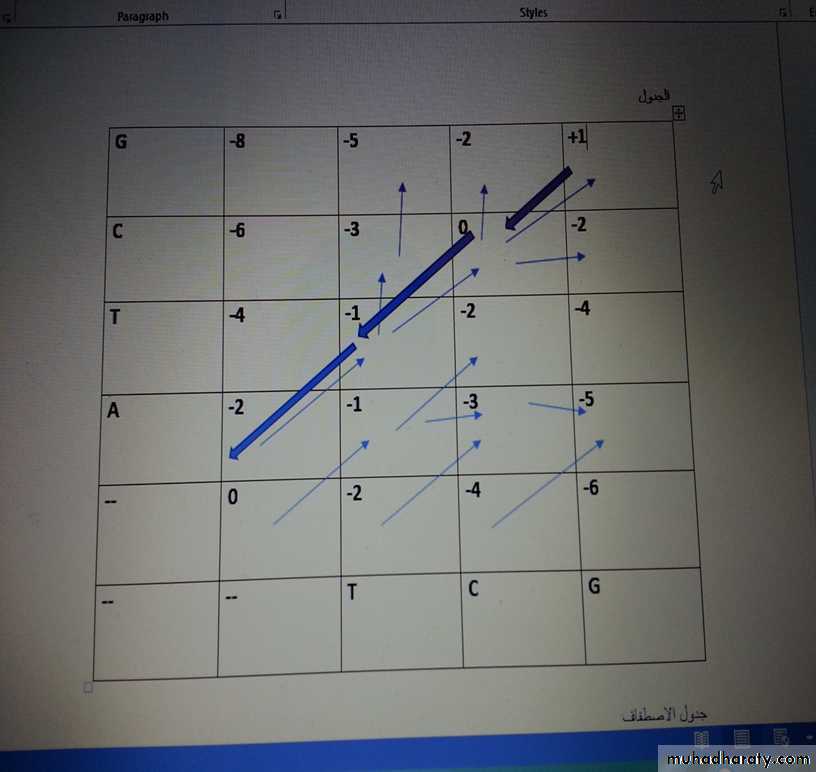
96
5- بعد اكمال كل المربعات نعمل رجوع اثري Trace Back ابتداء من اعلى قيمة ظهرت بالجدول (وهنا +1 ) , ومن هذه القيمة نعمل رجوع حسب الاسهم المؤشرة مسبقا ولكن باتجاه عكسي وكما في الجدول:
6- الخطوة الاخيرة والمهمة هي عمل الاصطفاف Alignment وكما يلي :
من اعلى قيمة نعمل ارجاع اثري باتجاه الصفر, وهنا اعلى قيمة +1 ومنها نعمل ارجاع وحسب الاسهم التي قمنا بتاشيرها , حيث نرجع صفر ثم -1 ثم -2 ونؤشر الرجوع بلون اخر (احمر),
بعد ذلك نعمل اصطفاف : عند الرقم +1 الرجع الاثري محوري , اذن نقارن بين الاحرف : هل مطابقة ام غير مطابقة G مع G , في هذا المثال مطابقة اذن نكتب الحرف العمودي بالاعلى والحرف الافقي بالاسفل ثم نعطي قيمة التطابق (ليس من الجدول) ولكن من القيمة المعطاة في السؤال وهي +1
المربع المحوري الاخر وقيمته صفر , ايضا مطابقة C مع C , اذن نكتب الحرفين بالاصطفاف, وايضا قيمته +1 لانها مطابقة
المربع المحوري الاخر, ايضا تطابق للحرف T مع T , اذن نكتب الحرفين بالاصطفاف, ونكتب قيمته +1 ايضا
.
97
98
المربع الاخير (وقيمته على الجدول -2 ) ويقابله الحرف A عموديا ولكن افقيا يقابله فجوى , اذن نكتب بالاصطفاف A بالاعلى , وبالاسفل نكتب فجوى والقيمة هنا -2 نكتبها بالاسفل زكما موضح ادناه وبذلك نكون قد انجزنا الاصطفاف,
وللتاكد بان عملنا صحيح : نكتب قيمة كل حرفين متقابلين , ثم نجمع فتكون النتيجة نفس قيمة اعلى مربع وهذا يدل على ان عملنا صحيح , اذا لم نحصل على نفس النتيجة معنى ذلك وجود خطا ما في احدى خطواتنا فيجب الاعادة.
C G T A
C G T -
+1 +1 +1 -2 , النتيجة هنا +1 وهي نفس قيمة اعلى قيمة في الجدول , اذن عملنا صحيح
الاصطفاف الموضعي Local Alignment
في هذا النوع من الاصطفاف تختلف بعض الخطوات عن الاصطفاف الشامل وكما يلي :مثال : اعمل اصطفاف للتوالي ادناه علما ان قيم التسجيل هي : تطابق =+1 و غير مطابق= -1, فجوة = -1
ATCG
TCG
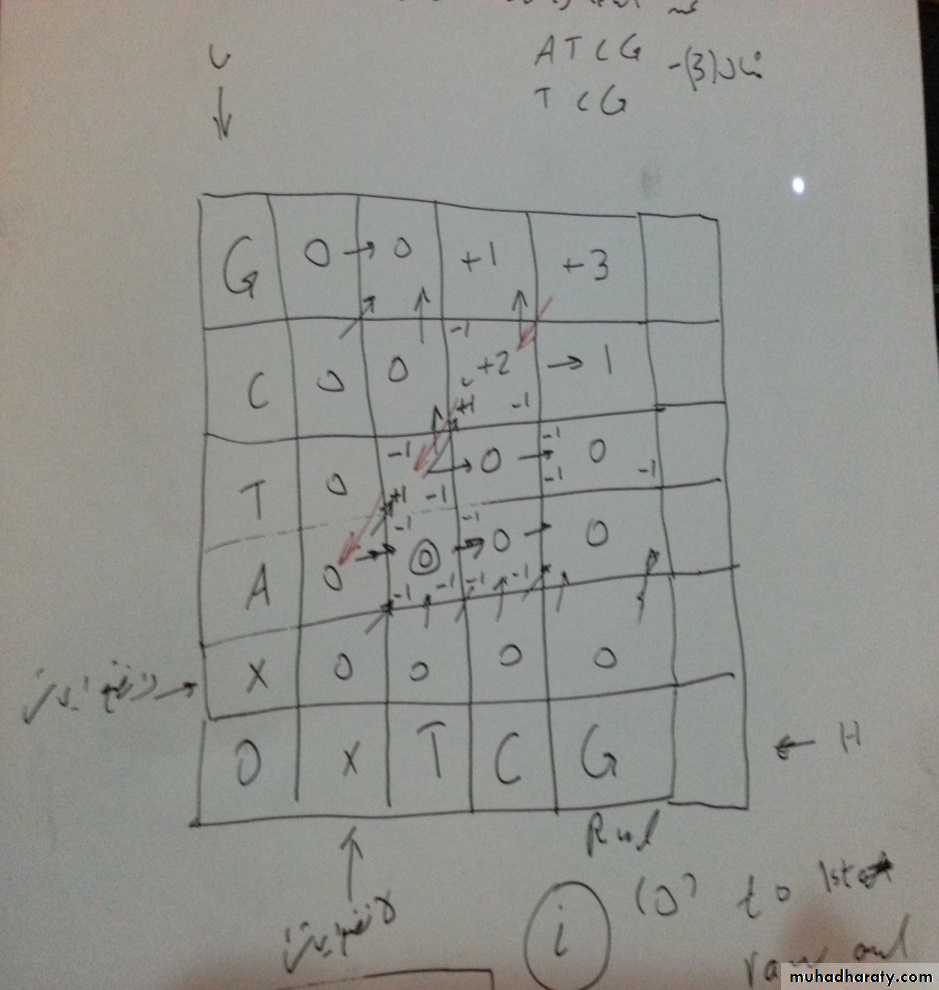
الخطوة االثانية : نضع القيم وبنفس الطريقة السابقة في الاصطفاف الشامل (نحسب قيمة كل مربع من ثلاث مربعات)
الخطوة الثالثة: نحدد اعلى قيمة بالجدول وهي هنا +3 , ثم نعمل ارجاع اثري الى الصفر
99
الخطوة الاولى : نرسم جدول ونضع التوالي العلوي في العمود الاول
ثم نضع التوالي السفلي بالصف السفلي من الجدول وكما في الجدول ادناه
نضع قيمة صفر في العمود والصف الاوليان فوق الاحرف
الخطوة الرابعة : نعمل اصطفاف ابتدا من اعلى قيمة (+3) حيث يقابلها نفس الحرف من الجهتين G لذا نضع حرفين اعلى واسفل , ثم ننتقل الى المربع المحوري التالي وقيمته +2 ويقبل حرفين متطابقين ( C) لذا نصف الحرفين الى جانب الحرفين الاوليين, ونفس الشئ يحصل مع المربع التالي وفيه حرفان متطابقين هما (T) , والمربع الاخير غير متطابق (فيه A مع فجوة) ونصف الحرف والفجوة بمكانهما ثم نضع قيم كل حرفين ونجد النتيجة النهائية التي يجب ان تكون نفس قيمة اعلى قيمة على الجدول وهي + 3 , ويكون الاصطفاف كما يلي :
ATCG
--TCG
+3 +2 -1 -1
100
BLAST أداة بحث الاصطفاف الموضعي الأساسي
Basic local alignment search toolسنة 1990 وهو من النوع الموجه ومنذ ذلك Stephen Altschul طور البرنامج من قبل
الوقت أصبح واحدا" من أكثر البرامج شهرة لتحليل وصف التواليات وكان البديل الأسرع
بدون التضحية بالدقة. يستخدم البرنامج الموجه لاصطفاف توالي FASTA للبرنامج المناظر له
الاستعلام مع جميع التواليات في قاعدة البيانات، والهدف هو إيجاد أعلى درجات التطابق للقطع
او الأجزاء بدون فجوات بين التواليات المتقاربة. ووجود مثل هذه الأجزاء أكثر من حد معين
يشير الى التشابه بين زوج التواليات الذي يميز بين التواليات المتقاربة عن التواليات المتباعدة
في قواعد البيانات.
101
.
Phylogenetic relationships العلاقات التطورية
يعد تحديد العلاقات التطورية بين الأحياء أحد المهام الرئيسة في المعلوماتية الحيوية ويعتمد
تحليل التواليات للجزيئات الحيوية الأساس في معرفة عملية التطور ورسم أشجار التطور.
ويوحي التشابه او الابتعاد بين التواليات الحيوية
الى المدى الذي مرت به التواليات من تغيير وتطور فمثلا التوالي
الاتي ماهي مسافة التغيير في التوالي ال دي ان اي ادناه :
acctga ® agcta
يشير الى حصول حالة استبدال وبعدها حالة حذف
. c وفي حالة الحذف تم حذف c مكان g ففي حالة الاستبدال ، تم استبدال
(نقطة من استبدال واحد ونقطة من حذف) ,مساوية 2 Edit distance فعليه فان مسافة التغيير
وهكذا مع بقية الامثلة وكما يلي
جد مسافة التغيير في التواليات التالية
, هنا حذف واحد لقاعدة السايتوسين = اذن المسافة نقطة واحدة actga ---- atga
هنا حذف للسايتوسين + استبدال الكوانين مع الادنين اذن مسافة التغيير هي 2 accctgg---- acctga
) اذن مسافة التغيير 3 وهكذا --------a مع القاعدة c + استبدال القاعدة (tg هنا حذف قاعدتين ( اذن نقطتين ttggcca--- tgcaa
102
103
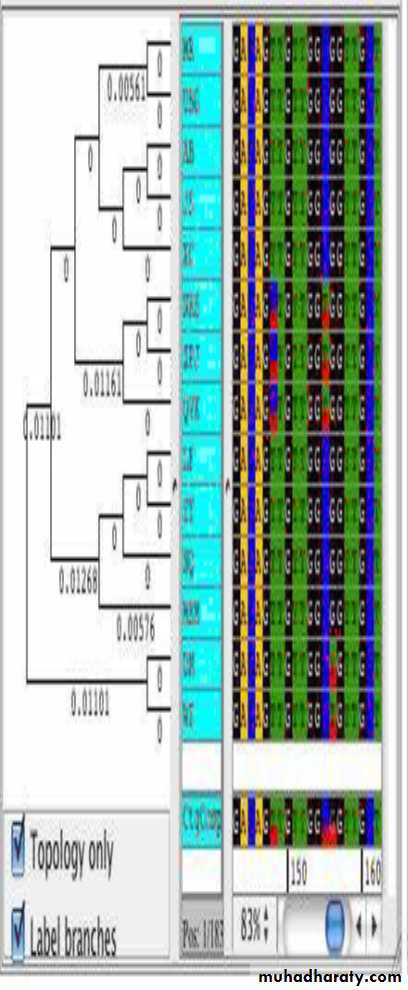
Phylogenetic relationships شكل يوضح العلاقات التطورية
التشعب يوضح مدى التقارب بين التواليات المختلفة ثم ربطها باصل واحد
104
Molecular clock فرضية الساعة الجزيئية
وهي تتضمن حدوث معدل ثابت من الطفرات في الكائن الحي تؤدي الى حصول تغيرات تطورية يمكن اعتمادها لمعرفة مدة الوقت المستغرق منذ اول حصول للطفرة للجينات.
كما ان التشابه العالي للتواليات يشير الى ان التغيير عن الأصل المشترك او الجد المشترك هو
جديد الحدوث ،
اما في حالة التشابه الواطئ فانه يشير الى انفراج وتباعد التواليات من زمن
والتي تشير الى حدوث معدل Molecular clock بعيد وهذا يعكس فرضية الساعة الجزيئية
ثابت من الطفرات وبالتالي التغيرات التطورية والتي تستخدم لاستنتاج الوقت المنقضي منذ
الانفراج الأول للجينات.
لذلك يمكن عد إيجاد العلاقات التطورية الجزيئية أحد مظاهر
المعلوماتية الحيوية المستعملة في التنبؤ بمدى حالة التطور الحاصلة للأحياء الموجودة اليوم
ايجاد التراكيب الخاصة في جينوم البكتريا
الصفات العامة للجينومات البكتريةحجوم البكتريا في العموم قد تكون ليست ذات علاقة مع محتواها من القواعد النتروجينية ولا
بالبيئة التي تعيش فيها الأحياء وتكون جينومات البكتريا والاركيا صغيرة نسبيا وتتراوح بين
95 % من الجينوم بين نصف الى 1 ميكا قاعدة، فضلا عن كون الكثافة الجينية عالية، فأكثر من 85
تحوي على تواليات مشفرة ولكن بعض الأحيان تصل الى 50 % وهذا يعني ان الكثافة
التشفيرية اعلى من حقيقة النواة
وعند المقارنة نجد ان التواليات المشفرة في الانسان مثلا قد لايتجاوز ال 30 %
Plandromic repeats ومن اهم التراكيب الخاصة تلك التي تدعى المكررات المقلوبة
105
المكررات المقلوبة Plandromic repeats في جينومات البكتريا
النقطة التي تؤخذ بنظر الاعتبار في جينومات البكتريا انها : تحوي القليل من المكررات مقارنة
بجينومات حقيقيات النواة واذا وجدت فهي بشكل : 1- نسخ متعددة لجين معين كما هو الحال مع
2- تكون بشكل جينات متناظرة نتجت من ،rRNA اوبرونات
تضاعف الجينات بدافع من قوى التطور، 3- بشكل مكررات في المناطق
(غير المشفرة مثل المكررات المقلوبة
وهذه توفر الحماية ضد غزو العاثيات (الفايروسات) palindromic repeats)
مثال لايجاد المكررات المقلوبة من اي تسلسل :
بين هل التتابع التالي يحتوي على مكررات مقلوبة Palindroms اذا يوجد بين طول القطع ومواقعهاالتتابع:
R= CGTAAGCTATATATAAT
8
TAATATATATGCAATGC
اذن يوجد مكرر واحد مقلوب بعدد 10 قواعد (باللون الاحمر)ابتدءا من القاعدة رقم 8
106