Membrane Potentials and
Action PotentialsElectrical potentials exist across the membranes of virtually all cells of the body. Some cells, such as nerve and muscle cells, are capable of generating rapidly changing electrochemical impulses at their membranes, and these impulses are used to transmit signals along the nerve or muscle membranes.
Nerve cell
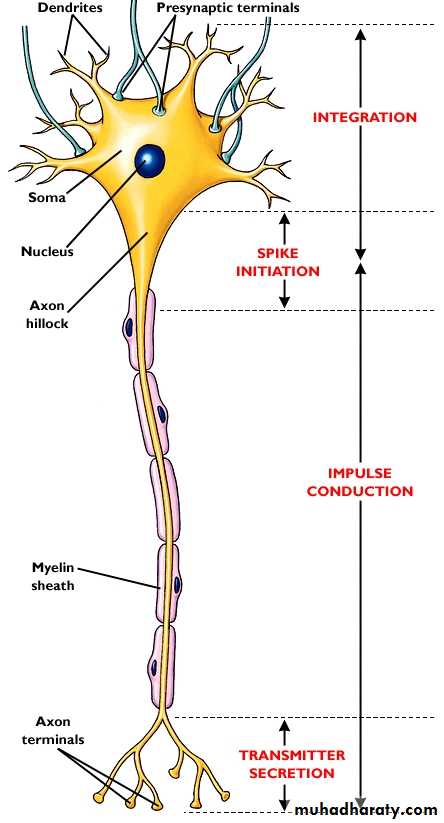
Represent the building block of the nervous system with specialized function of impulse transmission.
The nerve cell composed of cell body (soma) with 5-7 short projections (dendrites) and a longer fibrous axon.
The axon of some nerve fiber have a myelin sheath ,it is a protein insulator led down by Schwann cell which wrapping around the axon, the sheath envelops the axon except at its end and there are periodic constriction of 1 mm distance called node of ranvier so according to the present or absent of this sheath the nerve fiber can be :
myelinated nerve fiber .
un myelinated nerve fiber.
The axon ends in number of synaptic knobs (terminal buttons )which store the neuro-transmitter synthestized by the cell body.
Membrane potentials.
Separated electrical charges of opposite sign have the potential to do work if they are allowed to come together this potential is called an electrical potential. The unit of electrical potential are (volt) and (millivolt).
Membrane potentials caused by diffusion.
The concentration difference of ions; high Na+ in extra cellular fluid(ECF) and high K+ in intra cellular fluid (ICF); across a selectively permeable membrane can under appropriate conditions create a membrane potential.
Nerve cell
Resting membrane potential (RMP).
Electrical potential exists cross the membrane of essentially all body cells with a negative charge inside the cell relative to the positive charge outside. The resting membrane potential of large nerve fiber ,when not transmitting nerve signals is about (–90 millivolts).
The factors that determine the level of this resting potential :
1. Active transport of Na+ and K+ ions through the membrane (the Na-K pump) that all cell membranes of the body have a powerful Na-K pump that continually pumps sodium ions to the outside of the cell and potassium ions to the inside this is an electrogenic pump because more positive charges are pumped to the outside than to the inside (three Na+ ions to the outside for each two K+ ions to the inside), leaving a net deficit of positive ions on the inside; this causes a negative potential inside the cell membrane.2. Leakage of K+ and Na+ through the nerve membrane, in the nerve membrane there is a Protein channel where potassium and sodium ions can leak, called a potassium-sodium(K+ -Na+ ) “leak” channel. The channels are far more permeable to potassium than to sodium, normally about 100 times as permeable.
Nerve Action Potential:
Nerve signals are transmitted by action potentials, which are rapid changes in the membrane potential that spread rapidly along the nerve fiber membrane. The successive stages of the action potential are as follows.
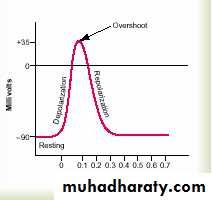
Resting Stage.
This is the resting membrane potential before the action potential begins. The membrane is said to be “polarized” during this stage because of the
(–90 millivolts) negative membrane potential that is present.
Depolarization Stage. At this time, the membrane suddenly becomes very permeable to sodium ions, allowing tremendous numbers of positively charged sodium ions to diffuse to the interior of the axon. The
normal “polarized” state of (–90 millivolts) is immediately neutralized and even the potential rising rapidly in the positive direction.
Action potential lasts for 1 millisecond in large myelinated
nerve fiber.Repolarization Stage.
Within a few 10,000ths of a second the sodium channels begin to close and the potassium channels open more than normal. Then, rapid diffusion of potassium ions to the exterior, re-establishes the normal negative resting membrane potential.
Voltage-Gated Sodium and Potassium Channels:
The necessary actor in causing both depolarization and repolarization of the nerve membrane during the action potential are:
Voltage-gated sodium channel.
Voltage-gated potassium channel.
Voltage-Gated Sodium Channel:
This channel has two gates—one near the outside of the channel called the activation gate, and another near the inside called the inactivation gate, in normal resting membrane (membrane potential is –90 millivolts.) the activation gate is closed, which prevents any entry of sodium ions to the interior of the fiber through these sodium channels.
Voltage-Gated Potassium Channel and Its Activation
During the resting state, the gate of the potassium channel is closed, and potassium ions are prevented from passing through this channel to the exterior. When the membrane potential rises from –90 millivolts toward zero, this voltage change causes a conformational opening of the gate and allows increased potassium diffusion outward through the channel. However, because of the slight delay in opening of the potassium channels, for the most part, they open just at the same time that the sodium channels are beginning to close because of inactivation. Thus, the decrease in sodium entry to the cell and the simultaneous increase in potassium exit from the cell combine to speed the repolarization process, leading to full recovery of the resting membrane potential within another few 10,000ths of a second.
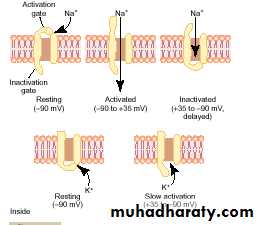
Initiation of the action potential
A Positive-Feedback Vicious Cycle Opens the Sodium Channels.First: as long as the membrane of the nerve fiber remains undisturbed, no action potential occurs in the normal nerve. However, if any event causes enough initial rise in the membrane potential from –90 millivolts toward the zero level, the rising voltage itself causes many voltage-gated sodium channels to begin opening. This allows rapid inflow of sodium ions, which causes a further rise in the membrane potential, thus opening still more voltage-gated sodium channels and allowing more streaming of sodium ions to the interior of the fiber. This process is a positive-feedback vicious cycle that, once the feedback is strong enough, continues until all the voltage-gated sodium channels have become activated (opened). Then, within another fraction of a millisecond, the rising membrane potential causes closure of the sodium channels as well as opening of potassium channels, and the action potential soon terminates.
Stimulus for nerve excitation :
1. chemical
2. mechanical
3. electrical.
The above factors increase Na+ permeability shifting the membrane potential toward the firing level causing an action potential.
Threshold for Initiation of the
Action PotentialAn action potential will not occur until the initial rise in membrane potential is great enough to create the vicious cycle. This occurs when the number of Na+ ions entering the fiber becomes greater than the number of K+ ions leaving the fiber. A sudden rise in membrane potential of 15 --30 millivolts usually is required. Therefore, a sudden increase in the membrane potential in a large nerve fiber from –90 millivolts up to about –65 millivolts usually causes the explosive development of an action potential. This level of –65 millivolts is said to be the threshold for stimulation.
Propagation of the action potential
An action potential elicited at any one point on an excitable membrane usually excites adjacent portions of the membrane, resulting in propagation of the action potential along the membrane.
The positive electrical charges are carried by the inward-diffusing sodium ions through the depolarized membrane and then for several millimeters in both directions along the core of the axon.
These positive charges increase the voltage for a distance of 1 to 3 millimeters inside the large myelinated fiber to above the threshold voltage value for initiating an action potential.
Therefore, the sodium channels in these new areas immediately open, and the explosive action potential spreads. These newly depolarized areas produce still more local circuits of current flow farther along the membrane, causing progressively more and more depolarization. Thus, the depolarization process travels along the entire length of the fiber. This transmission of the depolarization process along a nerve or muscle fiber is called a nerve or muscle impulse.
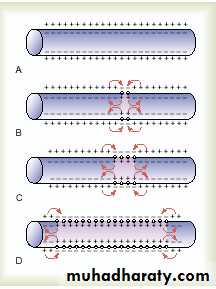
Propagation of the action potential in both side.
Direction of Propagation.An excitable membrane has no single direction of propagation, but the action potential travels in all directions away from the stimulus—even along all branches of a nerve fiber—until the entire membrane has become depolarized.
All-or-Nothing Principle.
Once an action potential has been elicited at any point on the membrane of a normal fiber, the depolarization process travels over the entire membrane if conditions are right, or it does not travel at all if conditions are not right. This is called the all-or-nothing principle, and it applies to all normal excitable tissues.
Therefore, for continued propagation of an impulse to occur, the ratio of action potential to threshold for excitation must at all times be greater than 1. This “greater than 1” requirement is called the safety factor for propagation.
Plateau in Some Action Potentials
In some instances, the excited membrane does not repolarize immediately after depolarization; instead, the potential remains on a plateau near the peak of the spike potential for many milliseconds, and only then does repolarization begin. The plateau greatly prolongs the period of depolarization. This type of action potential occurs in heart muscle fibers, where the plateau lasts for as long as 0.2 to 0.3 second and causes contraction of heart muscle to last for this same long period.
The cause of the plateau is a combination of several factors.
(1) voltage-activated sodium channels, called fast channels.
(2) voltage-activated calcium-sodium channels, which are slow to open and therefore are called slow channels.
Opening of fast channels causes the spike portion of the action potential, whereas the slow, prolonged opening of the slow calcium-sodium channels mainly allows calcium ions to enter the fiber, which is largely responsible for the plateau portion of the action potential as well.
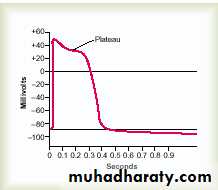
Plateau in Some Action Potentials
A second factor that may be partly responsible for the plateau is that the voltage-gated potassium channels are slower than usual to open, often not opening very much until the end of the plateau.This delays the return of the membrane potential toward its normal negative value of (–80 to –90 mv.).The role of Calcium Ions in action potential.
The membranes of almost all cells of the body have a calcium pump similar to the sodium pump, and calcium serves along with (or instead of) sodium in some cells to cause most of the action potential. Like the sodium pump, the calcium pump, pumps calcium ions from the interior to the exterior of the cell membrane (or into the endoplasmic reticulum of the cell) creating a calcium ion gradient of about 10,000-fold. The membrane of the cardiac and smooth muscle cells contain in addition, the voltage –gated Ca+ –Na+ (slow channels)which allow mainly Ca ion and to much less extend Na ions. when they open, both calcium and sodium ions flow to the interior of the fiber. The calcium channels are slow to become activated, requiring 10 to 20 times as long for activation as the sodium channels. Therefore, they are called slow channels, in contrast to the sodium channels,which are called fast channels. A decrease Ca ion concentration in extra cellular fluid (hypocalcaemia) lead to increase neuron excitability because of the profound effect of ECF Ca+ level on the ease of opening the voltage gated Na channels, when ECF Ca+ level falls 30-50% of its normal level ,it leads to spontaneous discharge in many peripheral nerves causing muscle tetany which could be lethal when involving respiratory muscle.
Special Characteristics of Signal Transmission in Nerve Trunks
The large fibers are myelinated, and the small ones are unmyelinated. The average nerve trunk contains about twice as many unmyelinated fibers as myelinated fibers. The central core of the fiber is the axon, and the membrane of the axon is the membrane that actually conducts the action potential. The axon is filled in its center with axoplasm, which is a viscid intracellular fluid. Surrounding the axon is a myelin sheath that is often much thicker than the axon itself. About once every 1 to 3 millimeters along the length of the myelin sheath is a node of Ranvier.
The myelin sheath is deposited around the axon by Schwann cells in the following manner: The membrane of a Schwann cell first envelops the axon; Then the Schwann cell rotates around the axon many times, laying down multiple layers of Schwann cell membrane containing the lipid substance sphingomyelin. This substance is an excellent electrical insulator that decreases ion flow through the membrane about 5000-fold. At the juncture between each two successive Schwann cells along the axon, a small uninsulated area only 2 to 3 micrometers in length remains where ions still can flow with ease through the axon membrane between the extracellular fluid and the intracellular fluid inside the axon. This area is called the node of Ranvier.
“Saltatory” Conduction in myelinated fibers from node to node.
Even though almost no ions can flow through the thick myelin sheaths of myelinated nerves, they can flow with ease through the nodes of Ranvier. Therefore, action potentials occur only at the nodes. Yet the action potentials are conducted from node to node, this is called saltatory conduction. That is electrical current flows through the surrounding extracellular fluid outside the myelin sheath as well as through the axoplasm inside the axon from node to node, exciting successive nodes one after another.Thus, the nerve impulse jumps down the fiber, which is the origin of the term “saltatory”.
Saltatory conduction is of value for two reasons.
First, by causing the depolarization process to jump long intervals along the axis of the nerve fiber, this mechanism increases the velocity of nerve transmission in myelinated fibers as much as 5- to 50-fold.
Second, saltatory conduction conserves energy for the axon because only the nodes depolarize, allowing perhaps 100 times less loss of ions than would otherwise be necessary.
Velocity of Conduction in Nerve Fibers.
The velocity of conduction in nerve fibers varies from as little as 0.25 m/sec in very small unmyelinated fibers to as great as 100 m/sec (the length of a football field in 1 second) in very large myelinated fibers.
Inhibition of Excitability—“Stabilizers” and Local Anesthetics
High extracellular fluid calcium ion concentration decreases membrane
permeability to sodium ions and simultaneously reduces excitability. Therefore, calcium ions are said to be a“stabilizer.”
Hypo K+: This decrease membrane potential to level which favors closure of most Na+ channels.
Local Anesthetics. including procaine and tetracaine. Most of these
act directly on the activation gates of the sodium channels, making it much more difficult for these gates to open, thereby reducing membrane excitability. When excitability has been reduced so low that the ratio of
action potential strength to excitability threshold (called the “safety factor”) is reduced below 1.0, nerve impulses fail to pass along the anesthetized nerves.
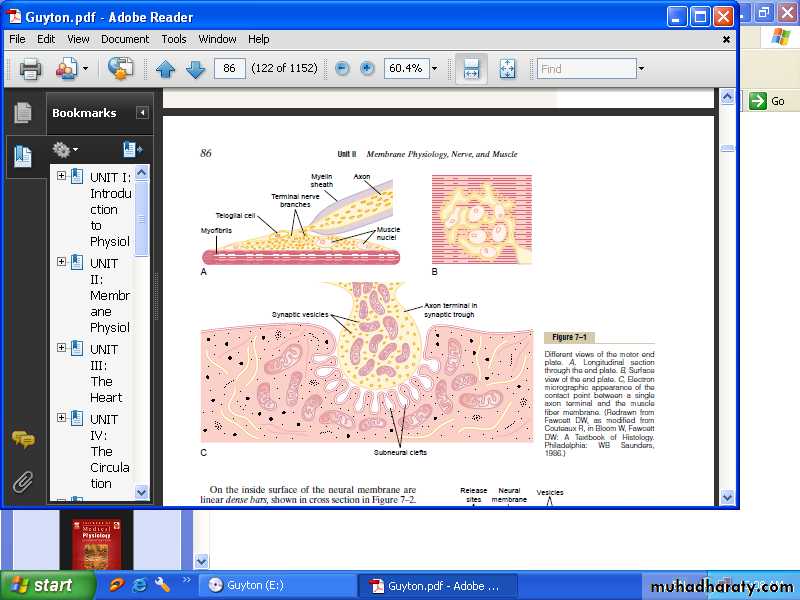
Physiologic anatomy of the neuromuscular junction- The motor end plate:
The nerve fiber forms a complex of branching nerve terminals that invaginate into the surface of the muscle fiber but lie outside the muscle fiber plasma membrane. The entire structure is called the motor end plate. The invaginated membrane is called the synaptic gutter or synaptic trough, and the space between the terminal and the fiber membrane is called the synaptic space or synaptic cleft. At the bottom of the gutter are numerous smaller folds of the muscle membrane called sub neural clefts, which greatly increase the surface area, at the nerve terminals there are many mitochondria that supply adenosine triphosphate (ATP), the energy source that used for synthesis of an excitatory transmitter acetylcholine. Acetylcholine(Ach) is synthesized in the cytoplasm of the nerve terminal. When a nerve impulse reaches the neuromuscular junction, Ach are released from the terminals into the synaptic space, In the synaptic space are large quantities of the enzyme acetyl cholinesterase, which destroys Ach a few milliseconds after it has been released from the synaptic vesicles. Ach stimulate calcium ions to diffuse from the synaptic space to the interior of the nerve terminal, calcium in turn stimulate more Ach vesicle release. Ach also stimulate the opening of acetylcholine-gated channels to allow large numbers of Na+ to pour inside the muscle fiber, carrying with them large numbers of positive charges. This creates a local positive potential change inside the muscle fiber membrane, called the end plate potential. Which in turn, initiates an action potential that spreads along the muscle membrane and thus causes muscle contraction.