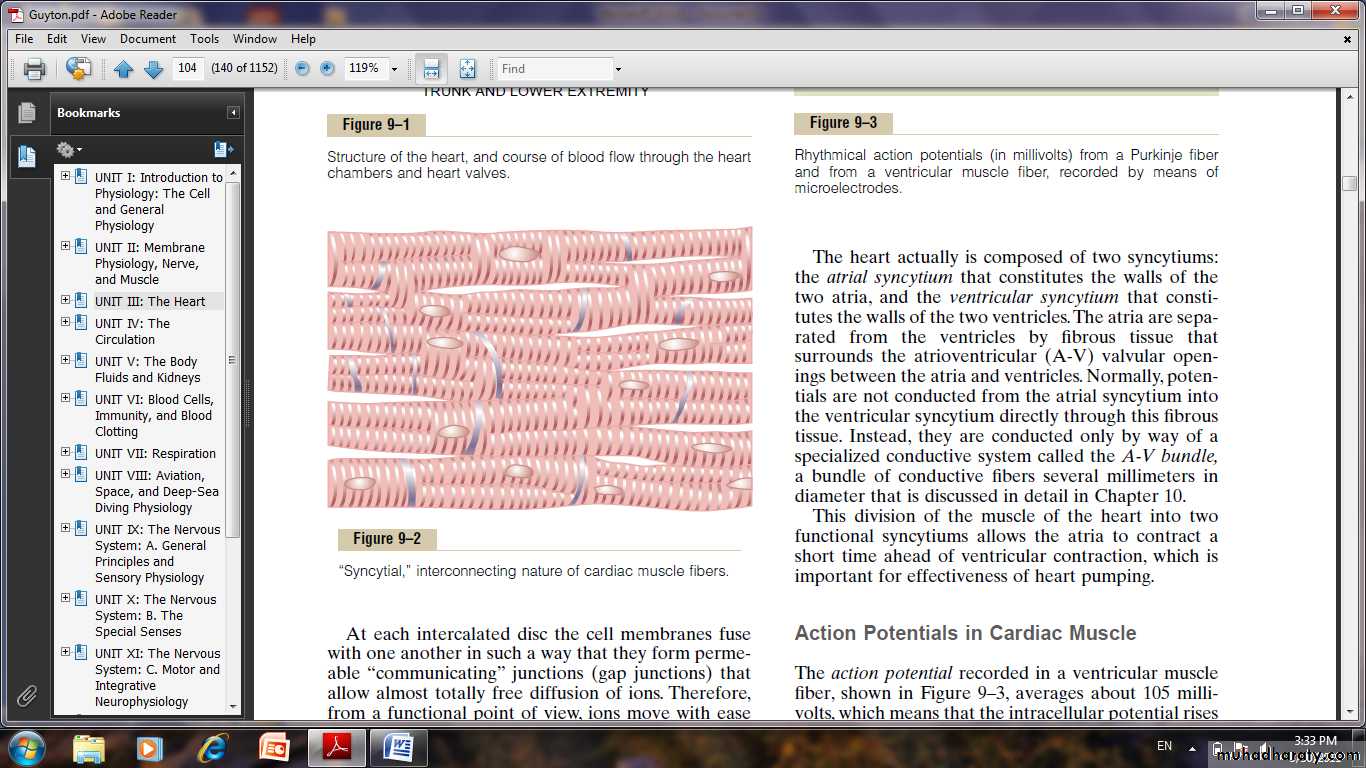
Heart Muscle
Physiology of Cardiac MuscleThe heart is composed of three major types of cardiac muscle: atrial muscle, ventricular muscle, and specialized excitatory and conductive muscle fibers. The atrial and ventricular types of muscle contract in much the same way as skeletal muscle, except that the duration of contraction is much longer.
Physiologic Anatomy of Cardiac Muscle
Cardiac muscle fibers arranged in a latticework, with the fibers dividing,
recombining, and then spreading again. The cardiac muscle is striated in the same manner as in typical skeletal muscle. Cardiac muscle has typical myofibrils that contain actin and myosin filaments almost identical to those found in skeletal muscle.
Cardiac Muscle as a Syncytium.
The dark areas crossing the cardiac muscle fibers are called intercalated discs; they are actually cell membranes that separate individual cardiac muscle cells from one another. That is, cardiac muscle fibers are made up of many individual cells connected in series and in parallel with one another. At each intercalated disc the cell membranes fuse with one another in such a way that they form permeable “communicating” junctions (gap junctions) that allow almost totally free diffusion of ions. So ions move with ease in the intracellular fluid along the longitudinal axes of the cardiac muscle fibers, so that action potentials travel easily from one cardiac muscle cell to the next, past the intercalated discs.
Thus, cardiac muscle is a syncytium of many heart muscle cells in which the cardiac cells are so interconnected that when one of these cells becomes excited, the action potential spreads to all of them, spreading from cell to cell throughout the latticework interconnections. The heart actually is composed of two syncytiums:
The atrial syncytium that constitutes the walls of the two atria, and the ventricular syncytium that constitutes the walls of the two ventricles. The atria are separated from the ventricles by fibrous tissue that surrounds the atrioventricular (A-V) valvular openings between the atria and ventricles. Normally, potentials are not conducted from the atrial syncytium into the ventricular syncytium directly through this fibrous tissue. Instead, they are conducted only by way of a specialized conductive system called the A-V bundle, a bundle of conductive fibers several millimeters in diameter.
This division of the muscle of the heart into two functional syncytiums allows the atria to contract a short time ahead of ventricular contraction, which is important for effectiveness of heart pumping.
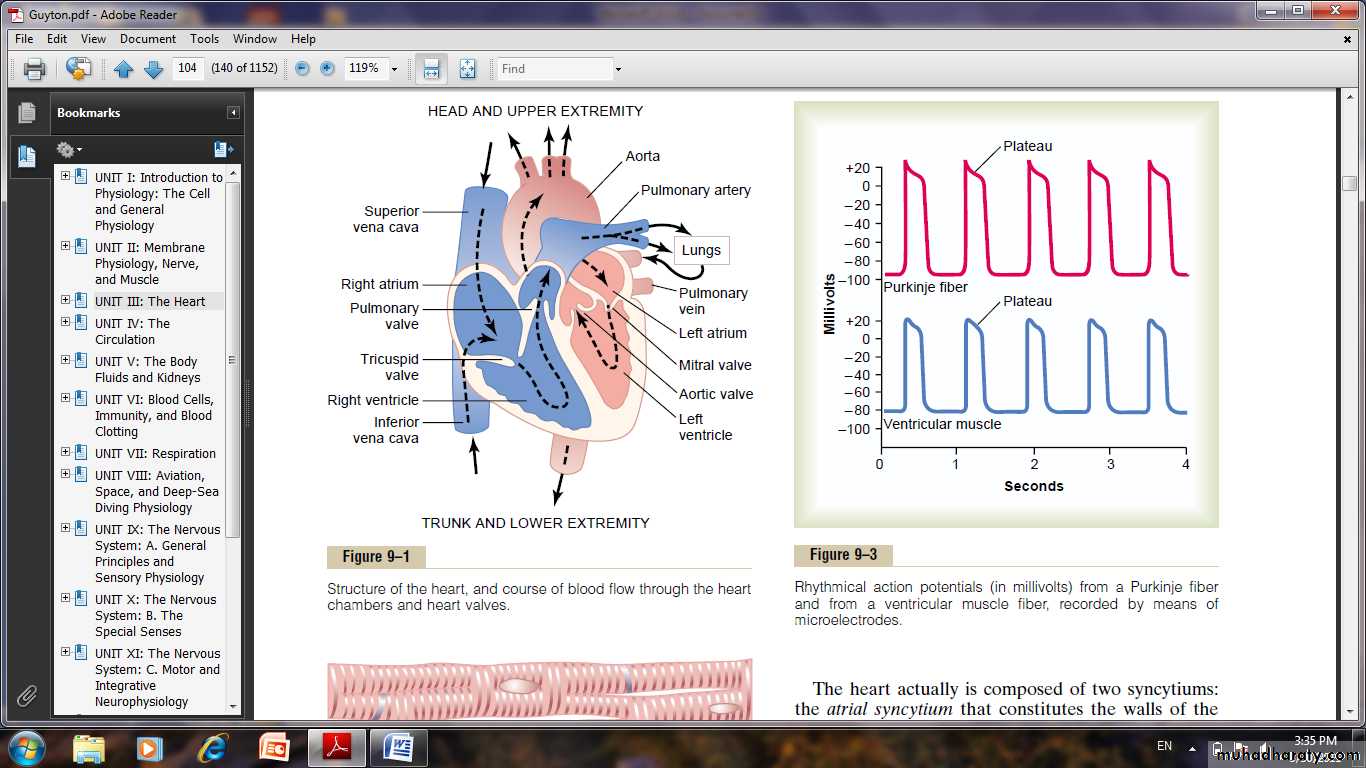
Action Potentials in Cardiac Muscle
The action potential recorded in a ventricular muscle fiber, averages about 105 millivolts, which means that the intracellular potential rises from a very negative value, about -85 millivolts, between beats to a slightly positive value, about +20 millivolts, during each beat. After the initial spike, the membrane remains depolarized for about 0.2 second, exhibiting a plateau, followed at the end of the plateau by abrupt repolarization. The presence of this plateau in the action potential causes ventricular contraction to last as much as 15 times as long in cardiac muscle as in skeletal muscle.
The Long Action Potential and the Plateau.
In cardiac muscle, the action potential is caused by opening of two types of channels:(1) The fast sodium channels as those in skeletal muscle .
(2) Slow calcium channels, which are also called calcium-sodium channels. This second population of channels differs from the fast sodium channels in that they are slower to open and, even more important, remain open for several tenths of a second. During this time, a large quantity of both calcium and sodium ions flows through these channels to the interior of the cardiac muscle fiber, and this maintains a prolonged period of
depolarization, causing the plateau in the action potential. Further, the calcium ions that enter during this plateau phase activate the muscle contractile process, while the calcium ions that cause skeletal muscle contraction are derived from the intracellular sarcoplasmic reticulum.
The second major functional difference between cardiac muscle and skeletal muscle that helps account for both the prolonged action potential and its plateau is this: Immediately after the onset of the action potential,
the permeability of the cardiac muscle membrane for potassium ions decreases about fivefold, an effect that does not occur in skeletal muscle. This decreased potassium permeability may result from the excess
calcium influx through the calcium channels just noted. Regardless of the cause, the decreased potassium permeability greatly decreases the outflux of positively charged potassium ions during the action potential plateau and thereby prevents early return of the action potential voltage to its resting level. When the slow calcium-sodium channels do close at the end of 0.2 to 0.3 second and the influx of calcium and sodium ions ceases, the membrane permeability for potassium ions also increases rapidly; this rapid loss of potassium from the fiber immediately returns the membrane potential to its resting level, thus ending the action potential.
Velocity of Signal Conduction in Cardiac Muscle.
The velocity of conduction of the excitatory action potential signal along both atrial and ventricular muscle fibers is about 0.3 to 0.5 m/sec, or about 1/250 the velocity in very large nerve fibers and about 1/10 the velocity in skeletal muscle fibers. In the Purkinje fibers is as great as 4 m/sec.Refractory Period of Cardiac Muscle.
Cardiac muscle, like all excitable tissue, is refractory to restimulation during the action potential. Therefore, the refractory period of the heart is the interval of time, during which a normal cardiac impulse cannot re-excite an already excited area of cardiac muscle. The normal refractory period of the ventricle is 0.25 to 0.30 second, which is about the duration of the prolonged plateau action potential.
Excitation-Contraction Coupling—Function of Calcium Ions and the Transverse Tubules.
When an action potential passes over the cardiac muscle membrane, the action potential spreads to the interior of the cardiac muscle fiber along the membranes of the transverse (T) tubules. The T tubule action potentials in turn act on the membranes of the longitudinal sarcoplasmic tubules to cause release of calcium ions into the muscle sarcoplasm from the sarcoplasmic reticulum. These calcium ions promote sliding of the actin and myosin filaments along one another; this produces the muscle contraction.
A large quantity of extra calcium ions also diffuses into the sarcoplasm from the T tubules themselves at the time of the action potential. Indeed, without this extra calcium from the T tubules, the strength of cardiac muscle contraction would be reduced considerably because the sarcoplasmic reticulum of cardiac muscle is less well developed than that of skeletal muscle and does not store enough calcium to provide full contraction.
The strength of contraction of cardiac muscle depends to a great extent on the concentration of calcium ions in the extracellular fluids. At the end of the plateau of the cardiac action potential, the influx of calcium ions to the interior of the muscle fiber is suddenly cut off, and the calcium ions in the sarcoplasm are rapidly pumped back out of the muscle fibers into both the sarcoplasmic reticulum and the T tubule–extracellular fluid space. As a result, the contraction ceases until a new action potential comes along.