Chapter 54 P.673 Motor Functions of the Spinal Cord The Cord Reflexes
Organization of the spinal cord for motor functions:The cord gray matter is the integrative area for the cord reflexes.
Sensory signals enter the cord almost entirely through the sensory (posterior) roots. Then, every sensory signal travels to two separate destinations:
(1) One branch of the sensory nerve terminates almost immediately in the gray matter of the cord and elicits local segmental cord reflexes and other local effects.

(2) Another branch transmits signals to higher levels of the nervous system—to higher levels in the cord itself, to the brain stem, or even to the cerebral cortex.
The other neurons are of two types:
(1)anterior motor neurons.
(2) interneurons.
Anterior motor neurons:
Located in each segment of the anterior horns of the cord gray matter are called anterior motor neurons.
They give rise to the nerve fibers that leave the cord by way of the anterior roots and directly innervate the skeletal muscle fibers.
The neurons are of two types, alpha motor neurons and gamma motor neurons.
Alpha motor neurons:
The alpha motor neurons give rise to large type A alpha (Aὰ) motor nerve fibers, averaging 14 micrometers in diameter; these fibers branch many times after they enter the muscle and innervate the large skeletal muscle fibers.Stimulation of a single alpha nerve fiber excites from three to several hundred skeletal muscle fibers, which are collectively called the motor unit.
Gamma motor neurons:
They transmit impulses through much smaller type A gamma (Ag) motor nerve fibers, averaging 5 micrometers in diameter, which go to small, special skeletal muscle fibers called intrafusal fibers.These fibers constitute the middle of the muscle spindle, which helps control basic muscle “tone”.
Interneurons:
Interneurons are present in all areas of the cord gray matter.
They are small and highly excitable, often exhibiting spontaneous activity and capable of firing as rapidly as 1500 times per second.
They have many interconnections with one another, and many of them also synapse directly with the anterior motor neurons.
The interconnections among the inter neurons and anterior motor neurons are responsible for most of the integrative functions of the spinal cord.
Muscle sensory receptors-muscle spindles and Golgi tendon organs — and their roles in muscle control
There are two special types of sensory receptors
• Muscle spindles:
• Which are distributed throughout the belly of the muscle and send information to the nervous system about muscle length or rate of change of length.
•
• (2)Golgi tendon organ:
• Which are located in the muscle tendons and transmit information about tendon tension or rate of change of tension.
• The signals from these two receptors are almost entirely for the purpose of intrinsic muscle control.
• They operate almost completely at a subconscious level.
• They transmit tremendous amounts of information not only to the spinal cord but also to the cerebellum, cerebral cortex, to control muscle contraction.

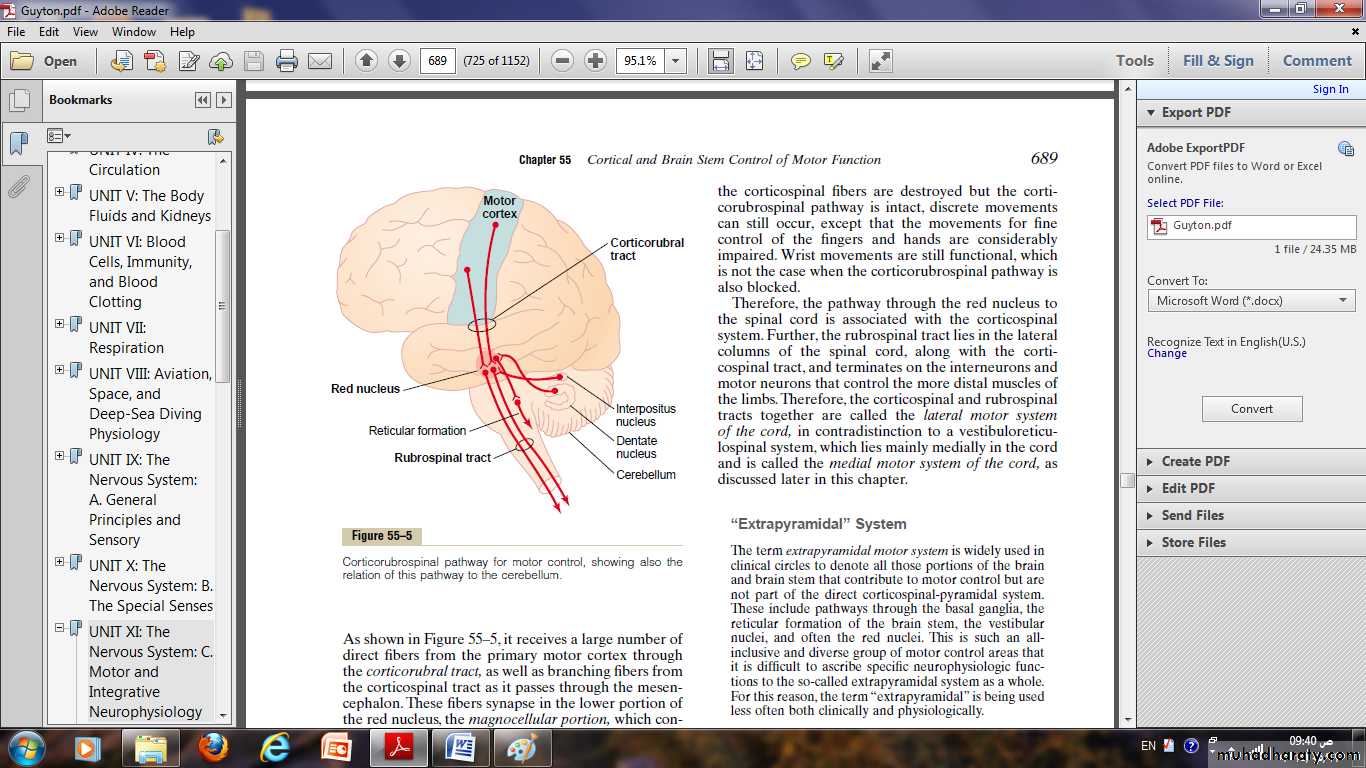
Receptor function of the muscle spindle structure and motor innervation of the muscle spindle:
Each muscle spindle is 3 to 10 millimeters long.
It is built around 3 to 12 very small intrafusal muscle fibers, pointed at their ends and attached to the surrounding large extrafusal skeletal muscle fibers.
Each intrafusal muscle fiber is a very small skeletal muscle fiber.
However, the central region of each of these fibers has few or no actin and myosin filaments.
This central portion does not contract when the ends do, it functions as a sensory receptor.
The end portions that do contract are excited by small gamma motor nerve fibers (gamma efferent) that originate from small type A gamma motor neurons in the anterior horns of the spinal cord.
The large alpha efferent fibers (type A alpha nerve fibers) innervate the extrafusal skeletal muscle.
Sensory innervations of the muscle spindle:
The receptor portion of the muscle spindle is its central portion.In this area, the intrafusal muscle fibers do not have myosin and actin contractile elements.
Sensory fibers originate in this area; They are stimulated by stretching of this mid portion of the spindle, the muscle spindle receptor can be excited in two ways:
1. Lengthening the whole muscle stretches the mid portion of the spindle and, therefore, excites the receptor.
2. Contraction of the end portions of the spindle’s intrafusal fibers stretches the mid portion of the spindle and therefore excites the receptor.
Two types of sensory endings are found in this central receptor area of the muscle spindle.
They are the primary ending and the secondary ending.
Primary Ending:
In the center of the receptor area a large sensory nerve fiber encircles the central portion of each intrafusal fiber, forming the so called primary ending or annulospiral ending.This nerve fiber is a type Ia fiber averaging 17 micrometers in diameter, and it transmits sensory signals to the spinal cord at a velocity of 70 to 120 m/sec.
Secondary Ending.
Usually one but sometimes two smaller sensory nerve fibers—type II fibers with an average diameter of 8 micrometers—innervate the receptor region on one or both sides of the primary ending.This sensory ending is called the secondary ending; it encircles the intrafusal fibers in the same way that the type Ia fiber does, but often it spreads like branches on a bush.
Division of the Intrafusal Fibers into Nuclear Bag and Nuclear Chain Fibers—Dynamic and Static Responses of the Muscle Spindle.
There are also two types of muscle spindle intrafusal fibers:
(1) nuclear bag muscle fibers (one to three in each spindle), in which several muscle fiber nuclei are congregated in expanded “bags” in the central portion of the receptor area.
(2) nuclear chain fibers (three to nine), which are about half as large in diameter and half as long as the nuclear bag fibers and have nuclei aligned in a chain throughout the receptor area.
The primary sensory nerve ending (the 17-micrometer sensory fiber) is excited by both the nuclear bag intrafusal fibers and the nuclear chain fibers.
Conversely, the secondary ending (the 8-micrometer sensory fiber) is usually excited only by nuclear chain fibers.
Response of Both the Primary and the Secondary Endings to the Length of the Receptor—“Static” Response.
When the receptor portion of the muscle Spindle is stretched slowly, the number of impulses transmitted from both the primary and the secondary endings increases almost directly in proportion to the degree of stretching, and the endings continue to transmit these impulses for several minutes.
This effect is called the static response of the spindle receptor, meaning simply that both the primary and secondary endings continue to transmit their signals for at least several minutes if the muscle spindle itself remains stretched.
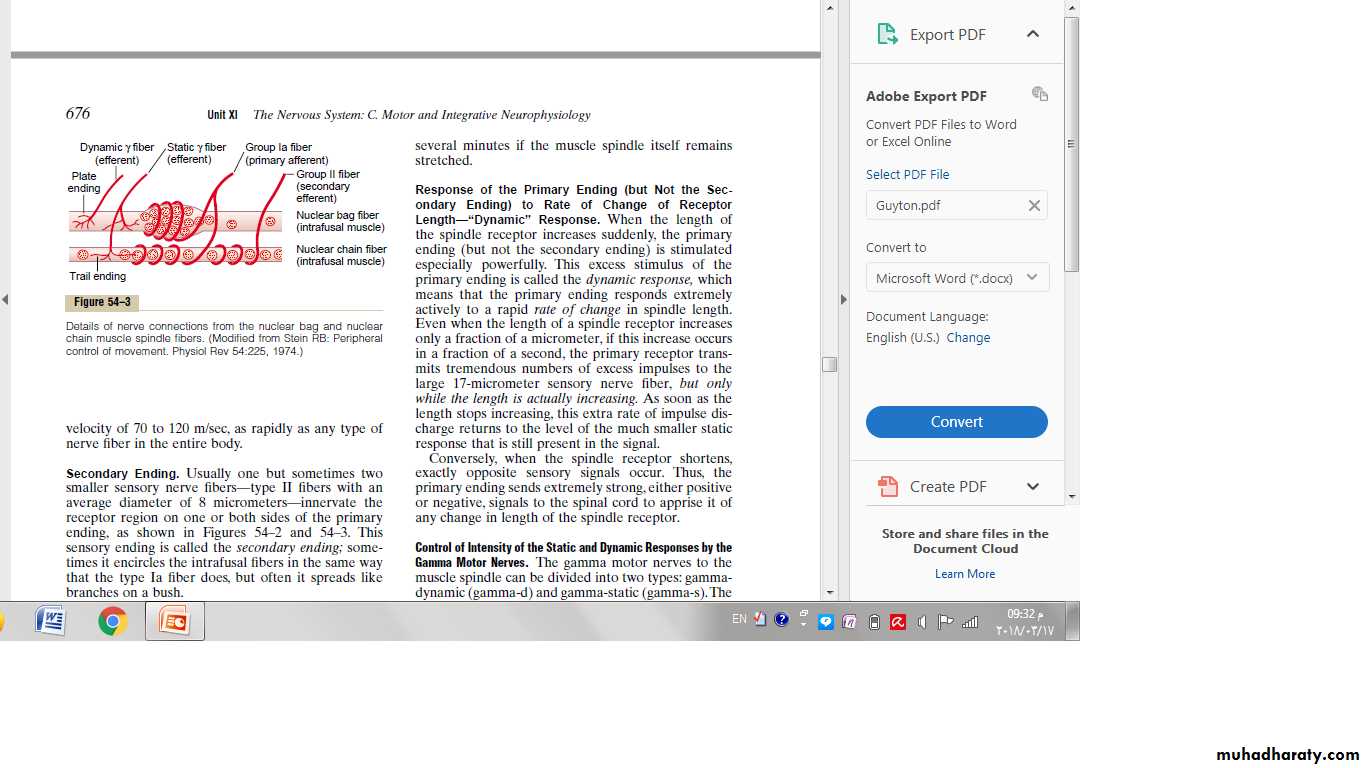
Response of the Primary Ending (but Not the Secondary Ending) to Rate of Change of Receptor Length—“Dynamic” Response.
When the length of the spindle receptor increases suddenly, the primary ending (but not the secondary ending) is stimulated especially powerfully.
This excess stimulus of the primary ending is called the dynamic response, which means that the primary ending responds extremely actively to a rapid rate of change in spindle length.
Even when the length of a spindle receptor increases only a fraction of a micrometer, if this increase occurs in a fraction of a second, the primary receptor transmits tremendous numbers of excess impulses to the large 17-micrometer sensory nerve fiber.
As soon as the length stops increasing, this extra rate of impulse discharge returns to the level of the much smaller static response that is still present in the signal.
Conversely, when the spindle receptor shortens, exactly opposite sensory signals occur.
Thus, the primary ending sends extremely strong, either positive or negative, signals to the spinal cord to apprise it of any change in length of the spindle receptor.Control of Intensity of the Static and Dynamic Responses by the Gamma Motor Nerves.
The gamma motor nerves to the muscle spindle can be divided into two types: gamma dynamic (gamma-d) and gamma-static(gamma-s).The first of these excites mainly the nuclear bag intrafusal fibers, and the second excites mainly the nuclear chain intrafusal fibers.
When the gamma-d fibers excite the nuclear bag fibers, the dynamic response of the muscle spindle becomes tremendously enhanced, whereas the static response is hardly affected.
Conversely, stimulation of the gamma-s fibers, which excite the nuclear chain fibers, enhances the static response while having little influence on the dynamic response.
Continuous Discharge of the Muscle Spindles Under Normal Conditions
Normally, when there is some degree of gamma nerve excitation, the muscle spindles emit sensory nerve impulses continuously.Stretching the muscle spindles increases the rate of firing, whereas shortening the spindle decreases the rate of firing.
Thus, the spindles can send to the spinal cord either positive signals—that is, increased numbers of impulses to indicate stretch of a muscle—or negative signals—below-normal numbers of impulses to indicate that the muscle is unstretched.
Muscle stretch reflex
The simplest manifestation of muscle spindle function is the muscle stretch reflex.Whenever a muscle is stretched suddenly, excitation of the spindles causes reflex contraction of the large skeletal muscle fibers of the stretched muscle and also of closely allied synergistic muscles.
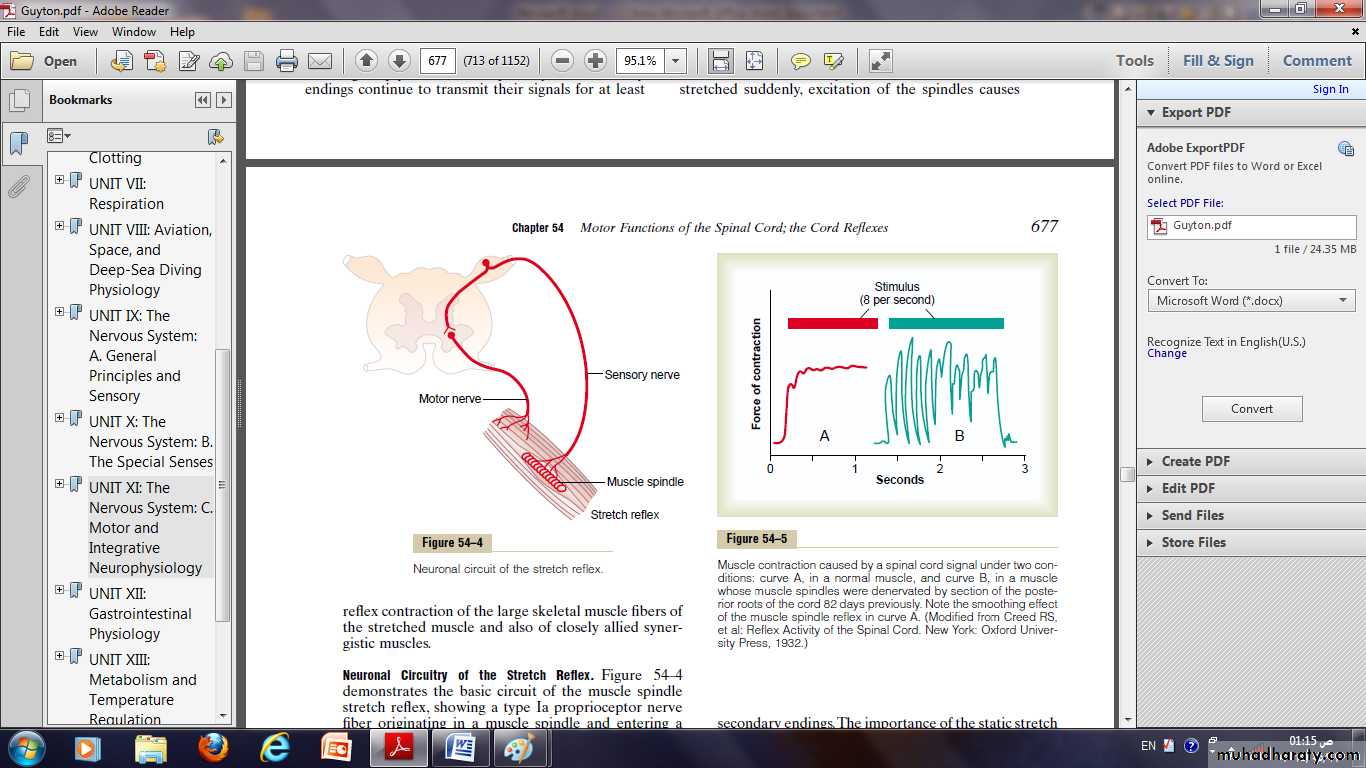
Neuronal circuitary of the stretch reflex:
The Ia proprioceptor nerve fiber originating in a muscle spindle and entering a dorsal root of the spinal cord.A branch of this fiber then goes directly to the anterior horn of the cord gray matter and synapses with anterior motor neurons that send motor nerve fibers back to the same muscle from which the muscle spindle fiber originated.
Thus, this is a monosynaptic pathway that allows a reflex signal to return with the shortest possible time delay back to the muscle after excitation of the spindle.
Dynamic Stretch Reflex and Static Stretch Reflexes.
The stretch reflex can be divided into two components: the dynamic stretch reflex and the static stretch reflex.The dynamic stretch reflex is elicited by the potent dynamic signal transmitted from the primary sensory endings of the muscle spindles, caused by rapid stretch or unstretch.
That is, when a muscle is suddenly stretched or unstretched, a strong signal is transmitted to the spinal cord; this causes an instantaneous strong reflex contraction (or decrease in contraction) of the same muscle from which the signal originated.
Thus, the reflex functions to oppose sudden changes in muscle length.
The dynamic stretch reflex is over within a fraction of a second after the muscle has been stretched (or unstretched) to its new length, but then a weaker static stretch reflex continues for a prolonged period thereafter.
This reflex is elicited by the continuous static receptor signals transmitted by both primary and secondary endings.
The importance of the static stretch reflex is that: it causes the degree of muscle contraction to remain reasonably constant.
Brain Areas for Control of the Gamma Motor System:
The gamma efferent system is excited specifically by signals from the:Bulbo-reticular facilitatory region of the brain stem and, secondarily, by impulses transmitted into the bulbo-reticular area from:
(1) The cerebellum.
(2) The basal ganglia.
(3) The cerebral cortex.
Muscle spindle system stabilizes body position during tense action
One of the most important functions of the muscle spindle system is to stabilize body position during tense motor action.To do this, the bulboreticular facilitatory region and its allied areas of the brain stem transmit excitatory signals through the gamma nerve fibers to the intrafusal muscle fibers of the muscle spindles.
This shortens the ends of the spindles and stretches the central receptor regions, thus increasing their signal output.
If the spindles on both sides of each joint are activated at the same time, reflex excitation of the skeletal muscles on both sides of the joint also increases, producing tight, tense muscles opposing each other at the joint.
The net effect is that: the position of the joint becomes strongly stabilized, and any force that tends to move the joint from its current position is opposed by highly sensitized stretch reflexes operating on both sides of the joint.
Clinical applications of the stretch reflex
Knee jerk and other muscle jerks:Clinically, a method used to determine the sensitivity of the stretch reflexes is to elicit the knee jerk.
The knee jerk can be elicited by simply striking the patellar tendon with a reflex hammer; this instantaneously stretches the quadriceps muscle and excites a dynamic stretch reflex that causes the lower leg to “jerk” forward.
Golgi tendon reflex: Golgi tendon organ helps control muscle tension:
Is an encapsulated sensory receptor through which muscle tendon fibers pass.About 10 to 15 muscle fibers are usually connected to each Golgi tendon organ, and the organ is stimulated when this small bundle of muscle fibers is “tensed” by contracting or stretching the muscle.
Thus, the major difference in excitation of the Golgi tendon organ versus the muscle spindle is that:
The spindle detects muscle length and changes in muscle length.
whereas the tendon organ detects muscle tension as reflected by the tension in itself.
Thus, Golgi tendon organs provide the nervous system with instantaneous information on the degree of tension in each small segment of each muscle.
The tendon organ, like the primary receptor of the muscle spindle.Has both:
A dynamic response.A static response.
Responding intensely when the muscle tension suddenly increases (the dynamic response).
But settling down within a fraction of a second to a lower level of steady-state firing that is almost directly proportional to the muscle tension (the static response).
Transmission of Impulses from the Tendon Organ into the central nervous system:
Signals from the tendon organ: are transmitted through large, rapidly conducting type Ib nerve fibers that average 16 micrometers in diameter.These fibers transmit signals both into local areas of the cord.
After synapsing in a dorsal horn of the cord through long fiber pathways such as the spinocerebellar tracts into the cerebellum and through still other tracts to the cerebral cortex.
The local cord signal excites a single inhibitory interneuron that inhibits the anterior motor neuron.
This local circuit directly inhibits the individual muscle without affecting adjacent muscles.
Inhibitory Nature of the Tendon Reflex and Its Importance:
When the Golgi tendon organs of a muscle tendon are stimulated by increased tension in the connecting muscle, signals are transmitted to the spinal cord to cause reflex effects in the respective muscle.This reflex is entirely inhibitory.
Thus, this reflex provides a negative feedback mechanism that prevents the development of too much tension on the muscle.
When tension on the muscle and on the tendon becomes extreme, the inhibitory effect from the tendon organ can be so great that it leads to a sudden reaction in the spinal cord that causes instantaneous relaxation of the entire muscle.
This effect is called the lengthening reaction; it is probably a protective mechanism to prevent tearing of the muscle or avulsion of the tendon from its attachments to the bone.
Flexor Reflex and the Withdrawal Reflexes
Almost any type of cutaneous sensory stimulus from a limb is likely to cause the flexor muscles of the limb to contract, thereby withdrawing the limb from the stimulating object.This is called the flexor reflex.
In its classic form, the flexor reflex is elicited most powerfully by stimulation of pain endings, such as by a pinprick, heat, or a wound, for which reason it is also called a nociceptive reflex, or simply a pain reflex.
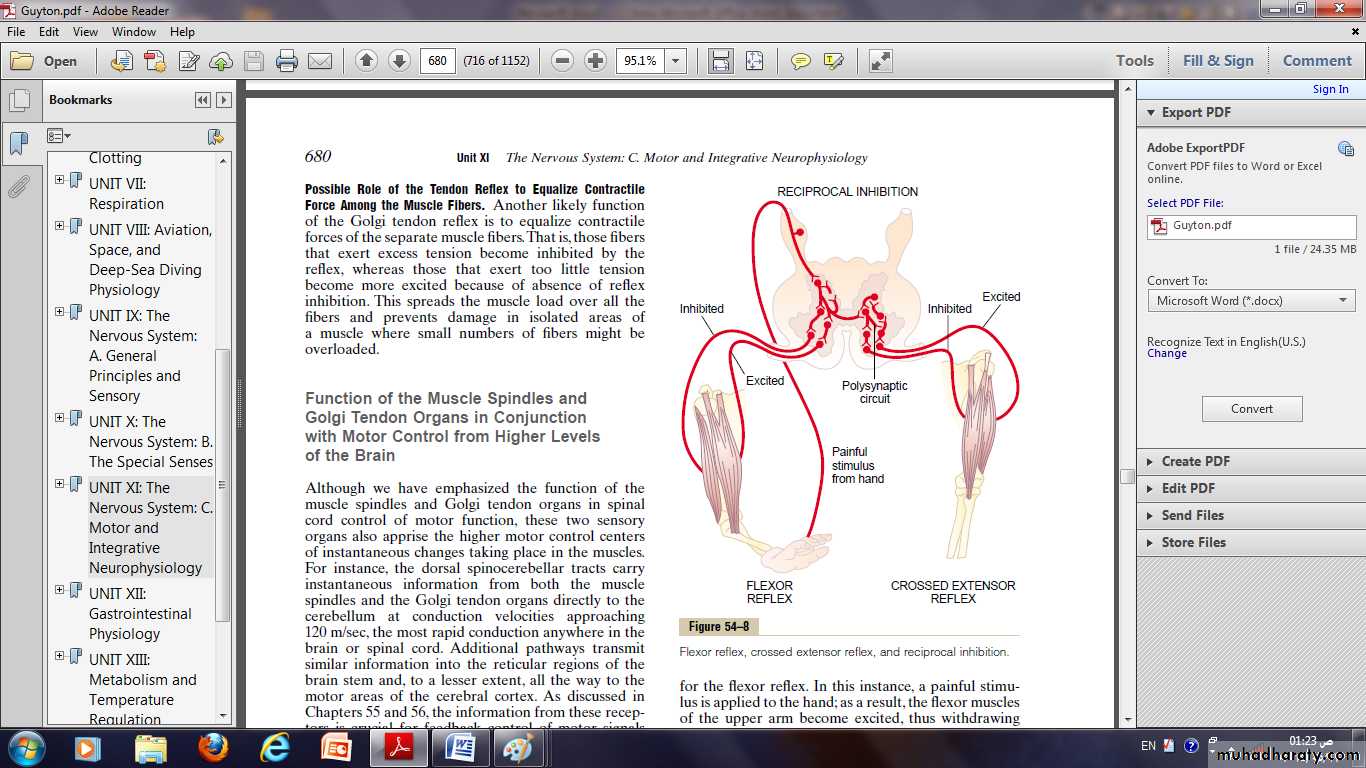
Stimulation of touch receptors can also elicit a weaker and less prolonged flexor reflex.
If some part of the body other than one of the limbs is painfully stimulated, that part will similarly be withdrawn from the stimulus, but the reflex may not be confined to flexor muscles, even though it is basically the same type of reflex.Therefore, the many patterns of these reflexes in the different areas of the body are called withdrawal reflexes.
Crossed Extensor Reflex
About 0.2 to 0.5 second after a stimulus elicits a flexor reflex in one limb, the opposite limb begins to extend.This is called the crossed extensor reflex.
Extension of the opposite limb can push the entire body away from the object causing the painful stimulus in the withdrawn limb.
Reciprocal Inhibition and Reciprocal Innervation
When a stretch reflex excites one muscle, is often simultaneously inhibits the antagonist muscles.This is the phenomenon of reciprocal inhibition, and the neuronal circuit that causes this reciprocal relation is called reciprocal innervation.
Chapter 55 685 Cortical and Brain Stem Control of Motor Function
Most “voluntary” movements initiated by the cerebral cortex are achieved when the cortex activates “patterns” of function stored in lower brain areas—the cord, brain stem, basal ganglia, and cerebellum.
These lower centers, in turn, send specific control signals to the muscles for a few types of movements.
The cortex has almost a direct pathway to the anterior motor neurons of the cord, bypassing some motor centers on the way.
This is especially true for control of the fine dexterous movements of the fingers and hands.
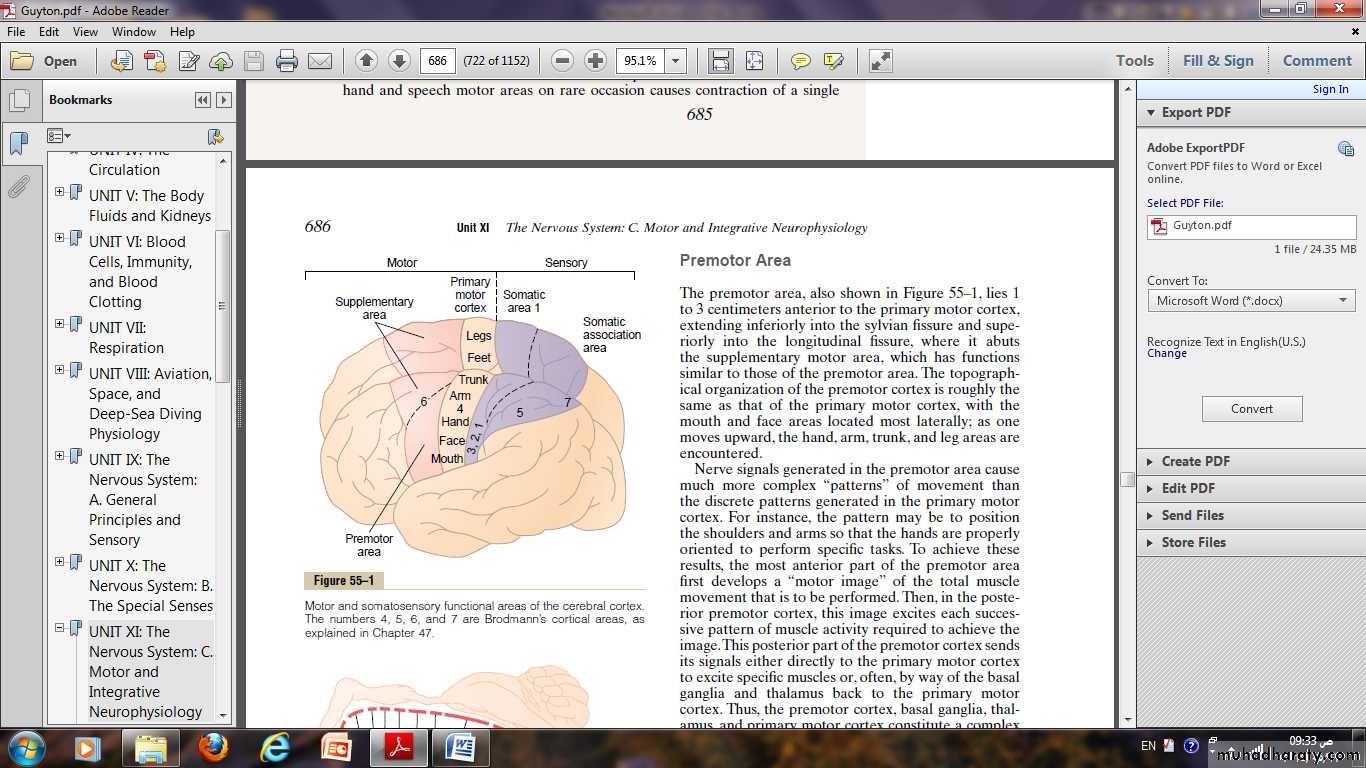
Motor cortex
It located anterior to the central cortical sulcus, occupying approximately the posterior one third of the frontal lobes.Posterior to the central sulcus is the somatosensory cortex which feeds the motor cortex by many of signals that initiate motor activities.
The motor cortex itself is divided into three subareas, each of which has its own topographical representation of muscle groups and specific motor functions:
(1) The primary motor cortex.
(2) The premotor area.
(3) The supplementary motor area.
Primary motor cortex:
It lies in the first convolution of the frontal lobes anterior to the central sulcu.The different muscle areas of the body are represented in the primary motor cortex.
Beginning with the face and mouth region near the sylvian fissure.
The arm and hand area, in the mid portion of the primary motor cortex.
The trunk, near the apex of the brain.
The leg and foot areas, in the part of the primary motor cortex that dips into the longitudinal fissure.

More than one half of the entire primary motor cortex is concerned with controlling the muscles of the hands and the muscles of speech.
So excitation of a single motor cortex neuron usually excites a specific movement rather than one specific muscle.
To do this, it excites a “pattern” of separate muscles, each of which contributes its own direction and strength of muscle movement.
Premotor Area:
The premotor area, lies 1 to 3 centimeters anterior to the primary motor cortex.Nerve signals generated in the premotor area cause much more complex “patterns” of movement.
The pattern may be to position the shoulders and arms so that the hands are properly oriented to perform specific tasks.
The premotor cortex in addition to basal ganglia, thalamus, and primary motor cortex constitute a complex overall system for the control of complex patterns of coordinated muscle activity.
Supplementary Motor Area
It lies mainly in the longitudinal fissure but extends a few centimeters onto the superior frontal cortex.Contractions elicited by stimulating this area are often bilateral rather than unilateral.
For instance, stimulation frequently leads to bilateral grasping movements of both hands simultaneously; these movements are perhaps rudiments of the hand functions required for climbing.
In general, this area functions in concert with the premotor area to provide:
Body-wide attitudinal movements.Fixation movements of the different segments of the body.
Positional movements of the head and eyes, and so forth.
Some specialized areas of motor control found in the human motor cortex:
Broca’s area and speech.It also called “word formation” it lies immediately anterior to the primary motor cortex and immediately above the sylvian fissure.
Damage to it does not prevent a person from vocalizing, but it does make it impossible for the person to speak whole words rather than un –coordinated utterances or an occasional simple word such as “no” or “yes.”
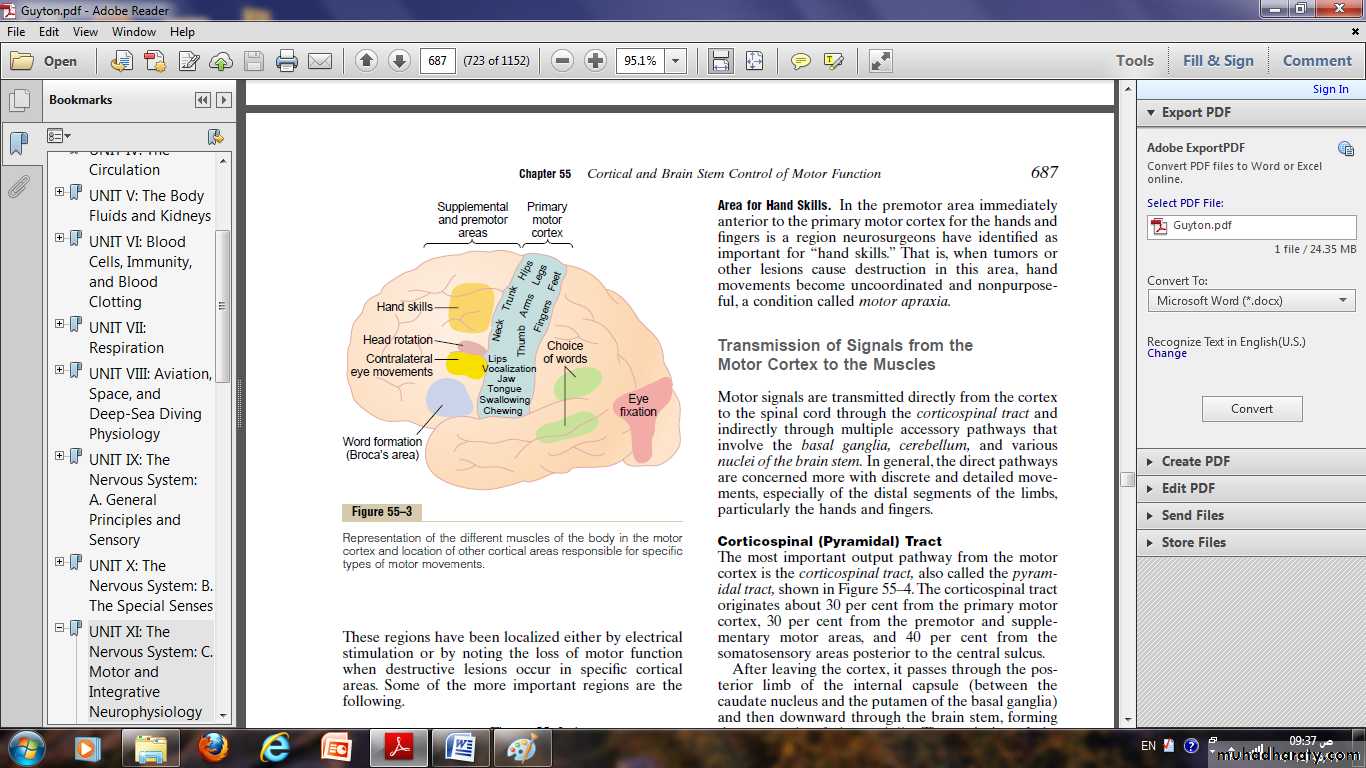
Sylvain fissure
“Voluntary” Eye Movement Field.In the premotor area immediately above Broca’s area is a locus for controlling voluntary eye movements.
Damage to this area prevents a person from voluntarily moving the eyes toward different objects.
Instead, the eyes tend to lock involuntarily onto specific objects.
This frontal area also controls eyelid movements such as blinking.
Head rotation area.
Slightly higher in the motor association area, electrical stimulation elicits head rotation.This area is closely associated with the eye movement field; it directs the head toward different objects.
Area for hand skills.
Immediately anterior to the primary motor cortex for the hands and fingers; important for “hand skills.”When tumors or other lesions destroy this area, hand movements become uncoordinated, non purposeful, a condition called motor apraxia.
Transmission of signals from the motor cortex to the muscles
Motor signals are transmitted directly from the cortex to the spinal cord through the corticospinal tract.And indirectly through multiple accessory pathways that involve the basal ganglia, cerebellum, and various nuclei of the brain stem.
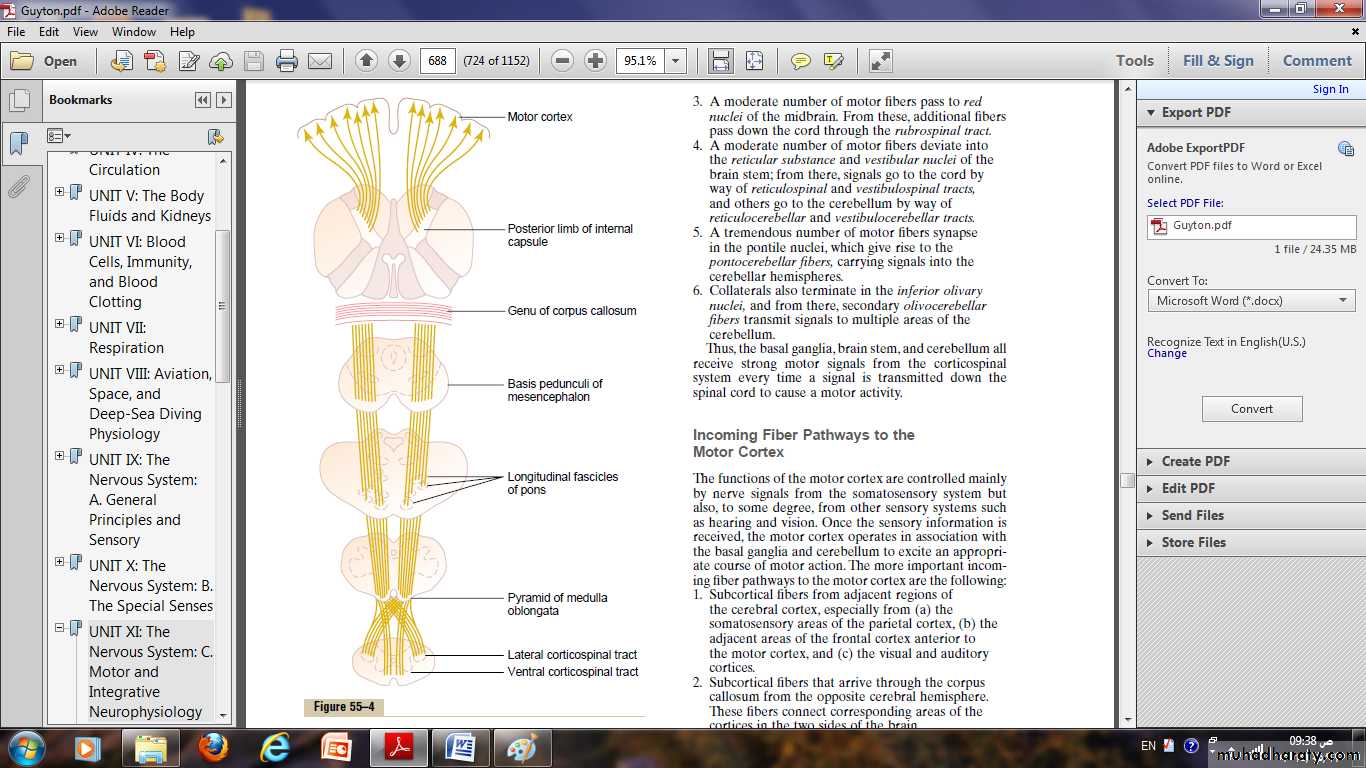
Corticospinal (Pyramidal) Tract
The most important output pathway from the motor cortex is the corticospinal tract, (pyramidal tract).The corticospinal tract originates about 30 per cent from the primary motor cortex, 30 per cent from the premotor and supplementary motor areas, and 40 per cent from the somato - sensory areas posterior to the central sulcus.
After leaving the cortex, it passes through the internal capsule and then downward through the brain stem.
The majority of the pyramidal fibers then cross in the lower medulla to the opposite side and descend into the lateral corticospinal tracts of the cord.
A few of the fibers do not cross to the opposite side in the medulla but pass ipsilaterally down the cord in the ventral corticospinal tracts.
finally terminating principally on the interneurons in the intermediate regions of the cord gray matter.
a few terminate on sensory relay neurons in the dorsal horn.
a very few terminate directly on the anterior motor neurons that cause muscle contraction.
The most impressive fibers in the pyramidal tract are a population of large myelinated fibers with a mean diameter of 16 micrometers.
“Extrapyramidal” System
It denote all those portions of the brain and brain stem that contribute to motor control but are not part of the direct corticospinal - pyramidal system.These include pathways through the basal ganglia, the reticular formation of the brain stem, the vestibular nuclei, and often the red nuclei.
Somatosensory feedback to the motor cortex helps control the precision of muscle contraction
When nerve signals from the motor cortex cause a muscle to contract, somatosensory signals return all the way from the activated region of the body to the neurons in the motor cortex that are initiating the action.
Most of these somatosensory signals arise in:
(1) The muscle spindles.(2)The tendon organs of the muscle tendons.
(3)The tactile receptors of the skin overlying the muscles.
These somatic signals often cause positive feedback enhancement of the muscle contraction in the following ways:
In the case of the muscle spindles, if the fusimotor muscle fibers in the spindles contract more than the large skeletal muscle fibers contract, the central portions of the spindles become stretched and, therefore, excited.
Signals from these spindles then return rapidly to the pyramidal cells in the motor cortex to signal them that the large muscle fibers have not contracted enough.
The pyramidal cells further excite the muscle, helping its contraction to catch up with the contraction of the muscle spindles.
Effect of lesions in the motor cortex or in the corticospinal pathway—the “Stroke”
This is caused either by a ruptured blood vessel that hemorrhages into the brain or by thrombosis of one of the major arteries supplying the brain.In either case, the result is loss of blood supply to the cortex or to the corticospinal tract.
Muscle spasticity caused by lesions that damage large areas adjacent to the motor cortex.
The primary motor cortex normally exerts a continual tonic stimulatory effect on the motor neurons of the spinal cord.when this stimulatory effect is removed, hypotonia results.
Most lesions of the motor cortex, especially those caused by a stroke, involve not only the primary motor cortex but also adjacent parts of the brain such as the basal ganglia.
So muscle spasm occurs in the afflicted muscle areas on the opposite side of the body (because the motor pathways cross to the opposite side).
This spasm results mainly from damage to accessory pathways from the non pyramidal portions of the motor cortex.
This is the spasticity that normally accompanies a “stroke” in a human being.
Role of the Brain Stem in Controlling Motor Function
The brain stem consists of the medulla, pons, and mesencephalon.It is an extension of the spinal cord upward into the cranial cavity, the brain stem is provides many special control functions, such as the following:
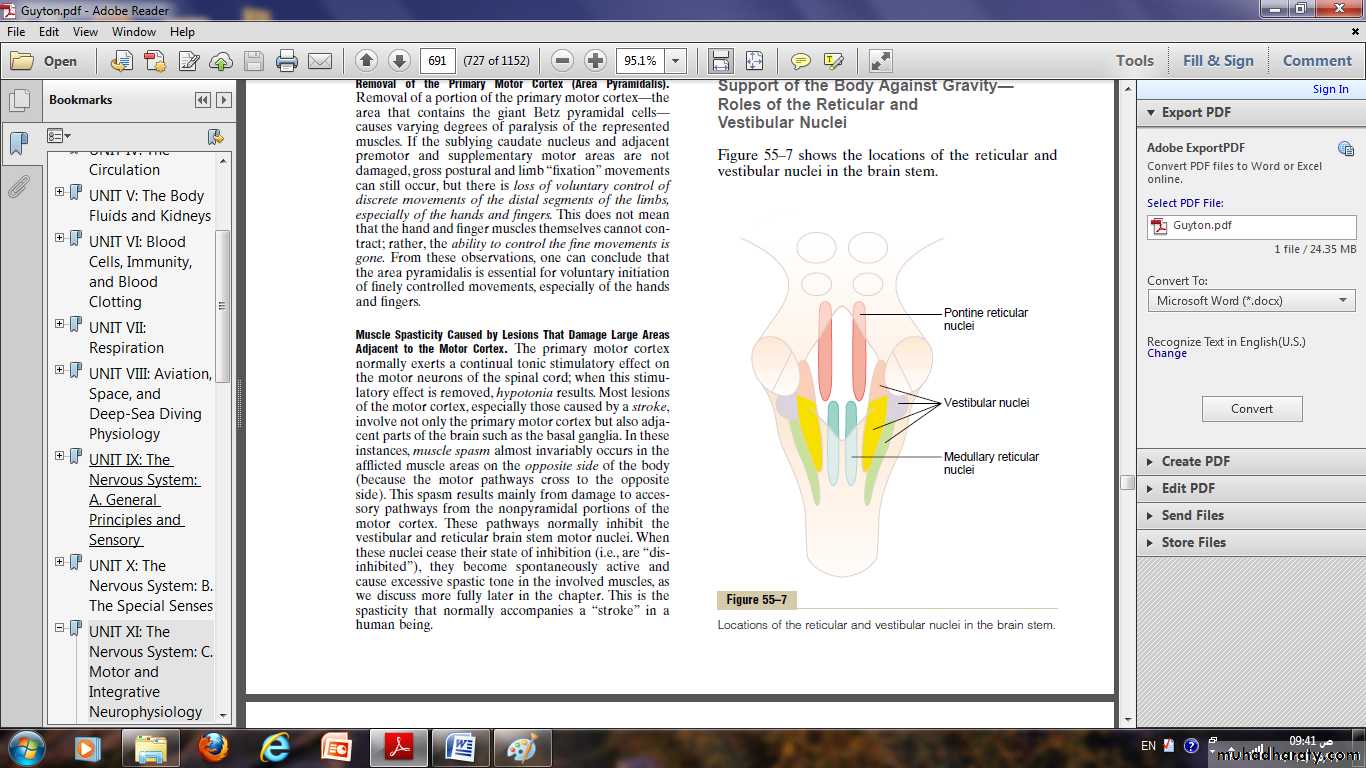
• Control of respiration.
• Control of the cardiovascular system.• Partial control of gastrointestinal function.
4. Control of many stereotyped movements of the body.
5. Control of equilibrium.
6. Control of eye movements.
Finally, the brain stem serves as a way station for “command signals” from higher neural centers.
GOOD LUCK