GIT system
Autonomic Control of the Gastrointestinal Tract
Objectives of our lecture:
• What is the effect of sympathetic and parasympathetic stimulation on GIT?• Hormonal control of GIT?
Parasympathetic Stimulation Increases Activity of the Enteric Nervous System.
The parasympathetic supply to the gut is divided into cranial and sacral divisions. Except for a few parasympathetic fibers to the mouth and pharyngeal regions of the alimentary tract, the cranial parasympathetic nerve fibers are almost entirely in the vagus nervesThese fibers provide extensive innervation to the esophagus, stomach, and pancreas and somewhat less to the intestines down through the first half of the large intestine. The sacral parasympathetics originate in the second, third, and fourth sacral segments of the spinal cord and pass through the pelvic nerves to the distal half of the large intestine and all the way to the anus.
The sigmoidal, rectal, and anal regions are considerably better supplied with parasympathetic fibers than are the other intestinal areas.
The postganglionic neurons of the gastrointestinal parasympathetic system are located mainly in the myenteric and submucosal plexuses.
Stimulation of these parasympathetic nerves causes general increase in activity of the entire enteric nervous system, which in turn enhances activity of most gastrointestinal functions.
Sympathetic Stimulation Usually Inhibits Gastrointestinal Tract Activity.
The sympathetic fibers to the gastrointestinal tract originate in the spinal cord between segments T5 and L2. Most of the preganglionic fibers that innervate the gut, after leaving the cord, enter the sympathetic chains that lie lateral to the spinal column, and many of these fibers then pass on through the chains to outlying ganglia such as to the celiac ganglion and various mesenteric gangliaMost of the postganglionic sympathetic neuron bodies are in these ganglia, and postganglionic fibers then spread through postganglionic sympathetic nerves to all parts of the gut.
The sympathetics innervate essentially all of the gastrointestinal tract, rather than being more extensive nearest the oral cavity and anus, as is true of the parasympathetics.
The sympathetic nerve endings secrete mainly norepinephrine. In general, stimulation of the sympathetic nervous system inhibits activity of the gastrointestinal tract, causing many effects opposite to those of the parasympathetic system. It exerts its effects in two ways: (1) to a slight extent by direct effect of secreted norepinephrine to inhibit intestinal tract smooth muscle (except the mucosal muscle, which it excites) and
(2) to a major extent by an inhibitory effect of norepinephrine on the neurons of the entire enteric nervous system. Strong stimulation of the sympathetic system can inhibit motor movements of the gut so greatly that this can literally block movement of food through the gastrointestinal tract
Afferent Sensory Nerve Fibers From the Gut
Many afferent sensory nerve fibers innervate the gut. Some of the nerve fibers have their cell bodies in the enteric nervous system and some have them in the dorsal root ganglia of the spinal cord. These sensory nerves can be stimulated by (1) irritation of the gut mucosa,(2) excessive distention of the gut, or (3) the presence of specific chemical substances in the gut. Signals transmitted through the fibers can then cause excitation or, under other conditions, inhibition of intestinal movements or intestinal secretion.
In addition, other sensory signals from the gut go all the way to multiple areas of the spinal cord and even to the brain stem. For example, 80 percent of the nerve fibers in the vagus nerves are afferent rather than efferent. These afferent fibers transmit sensory signals from the gastrointestinal tract into the brain medulla which, in turn, initiates vagal reflex signals that return to the gastrointestinal tract to control many of its functions.
Gastrointestinal Reflexes
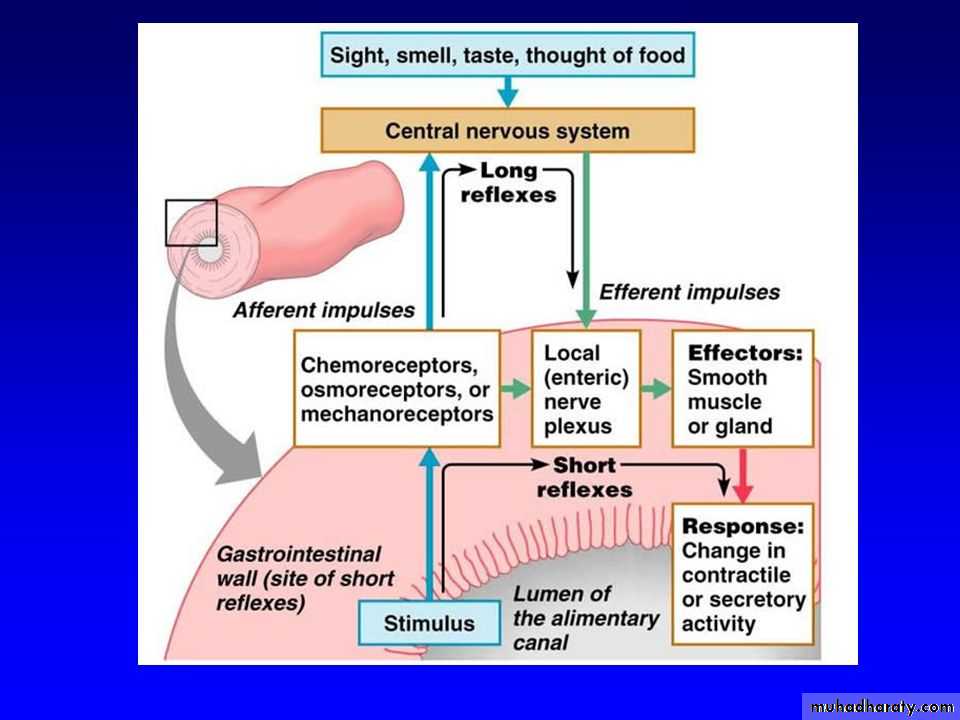
The anatomical arrangement of the enteric nervous system and its connections with the sympathetic and parasympathetic systems support three types of gastrointestinal reflexes that are essential to gastrointestinal control:1.Local reflexes:
Reflexes that are integrated entirely within the gut wall enteric nervous system. These reflexes include those that control much gastrointestinal secretion, peristalsis, mixing contractions, local inhibitory effects, and so forth2. Short reflexes:
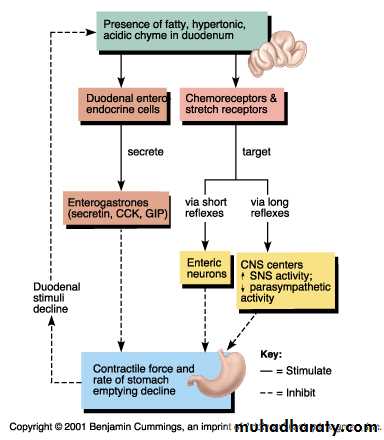
Reflexes from the gut to the prevertebral sympathetic ganglia and then back to the gastrointestinal tract. These reflexes transmit signals long distances to other areas of the gastrointestinal tract, such as signals from the stomach to cause evacuation of the colon (the gastrocolic reflex), signals from the colon and small intestine to inhibit stomach motility and stomach secretion (the enterogastric reflexes), and reflexes from the colon to inhibit emptying of ileal contents into the colon (the colonoileal reflex).3.Long reflexes:
Reflexes from the gut to the spinal cord or brain stem and then back to the gastrointestinal tract. These reflexes include especially (1) reflexes from the stomach and duodenum to the brain stem and back to the stomach—by way of the vagus nerves— to control gastric motor and secretory activity;(2) pain reflexes that cause general inhibition of the entire gastrointestinal tract; and (3) defecation reflexes that travel from the colon and rectum to the spinal cord and back again to produce the powerful colonic, rectal, and abdominal contractions required for defecation (the defecation reflexes)
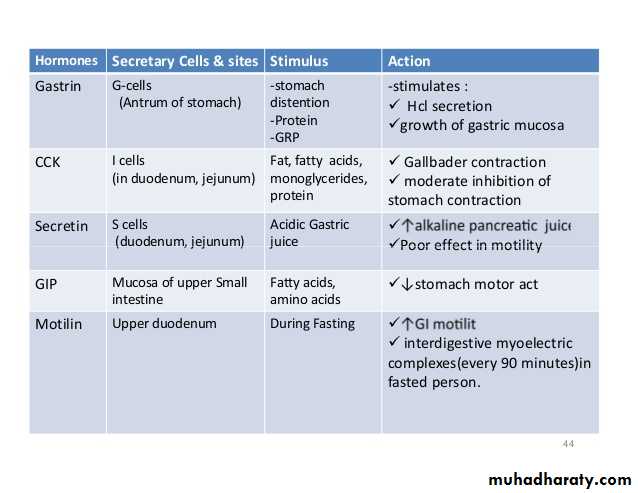
HORMONAL CONTROL OF GASTROINTESTINAL MOTILITY
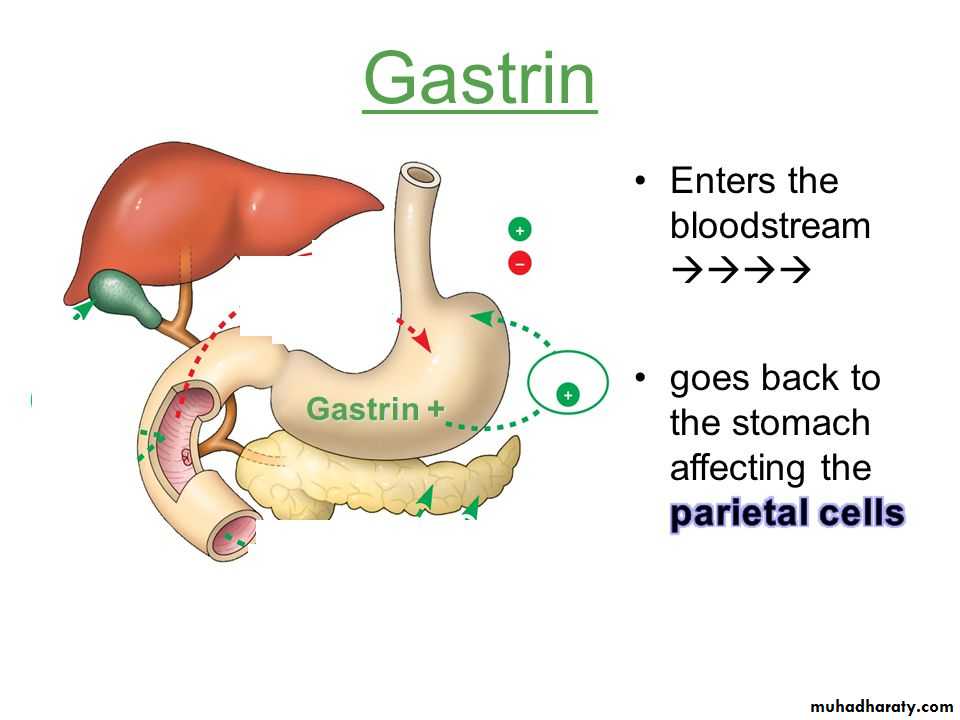
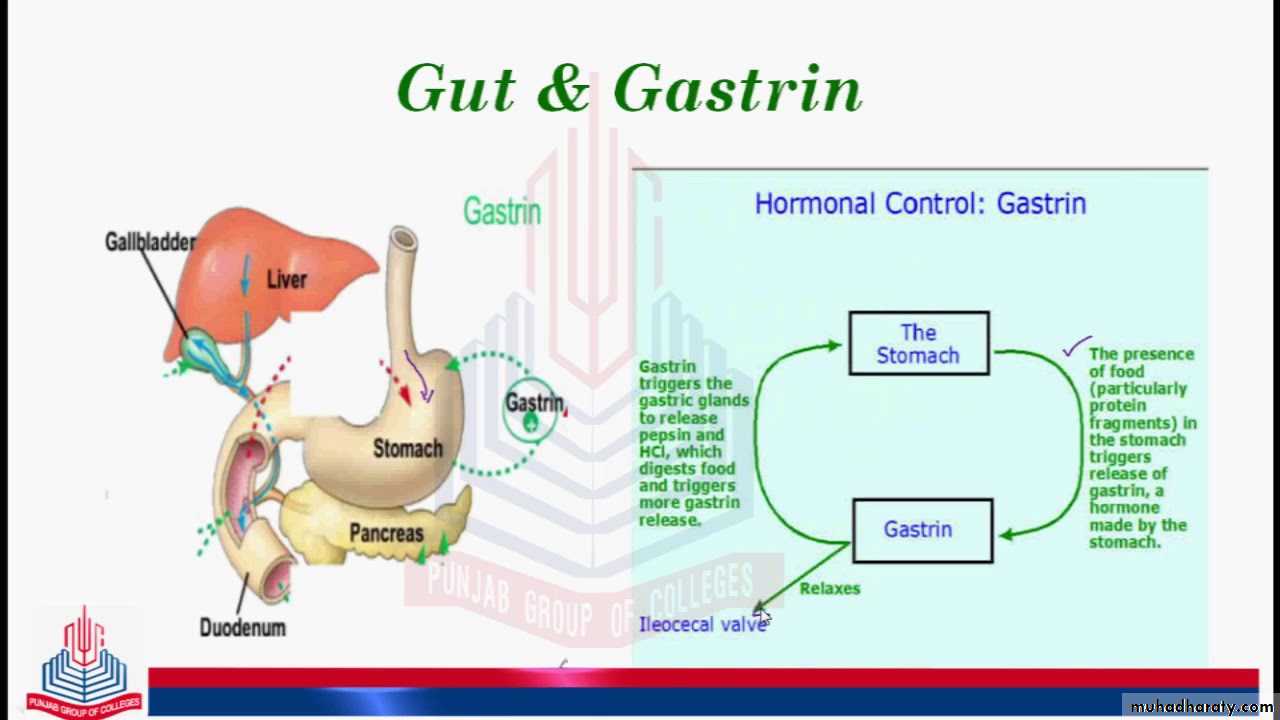
The gastrointestinal hormones are released into the portal circulation and exert physiological actions on target cells with specific receptors for the hormone. The effects of the hormones persist even after all nervous connections between the site of release and the site of action have been severedGastrin is secreted by the “G” cells of the antrum of the stomach in response to stimuli associated with ingestion of a meal, such as distention of the stomach, the products of proteins, and gastrin-releasing peptide, which is released by the nerves of the gastric mucosa during vagal stimulation. The primary actions of gastrin are (1) stimulation of gastric acid secretion and (2) stimulation of growth of the gastric mucosa.
Cholecystokinin (CCK) is secreted by “I” cells in the mucosa of the duodenum and jejunum mainly in response to digestive products of fat, fatty acids, and monoglycerides in the intestinal contents. This hormone strongly contracts the gallbladder, expelling bile into the small intestine, where the bile, in turn, plays important roles in emulsifying fatty substances and allowing them to be digested and absorbed.
CCK also inhibits stomach contraction moderately. Therefore, at the same time that this hormone causes emptying of the gallbladder, it also slows the emptying of food from the stomach to give adequate time for digestion of the fats in the upper intestinal tract. CCK also inhibits appetite to prevent overeating during meals by stimulating sensory afferent nerve fibers in the duodenum; these fibers, in turn, send signals by way of the vagus nerve to inhibit feeding centers in the brain .
Secretin, the first gastrointestinal hormone discovered, is secreted by the “S” cells in the mucosa of the duodenum in response to acidic gastric juice emptying into the duodenum from the pylorus of the stomach. Secretin has a mild effect on motility of the gastrointestinal tract and acts to promote pancreatic secretion of bicarbonate, which in turn helps to neutralize the acid in the small intestine
Glucose-dependent insulinotropic peptide (also called gastric inhibitory peptide [GIP]) is secreted by the mucosa of the upper small intestine, mainly in response to fatty acids and amino acids but to a lesser extent in response to carbohydrate. It has a mild effect in decreasing motor activity of the stomach and therefore slows emptying of gastric contents into the duodenum when the upper small intestine is already overloaded with food product Glucose-dependent insulinotropic peptide, at blood levels even lower than those needed to inhibit gastric motility, also stimulates insulin secretion.
Motilin is secreted by the stomach and upper duodenum during fasting, and the only known function of this hormone is to increase gastrointestinal motility. Motilin is released cyclically and stimulates waves of gastrointestinal motility called interdigestive myoelectric complexes that move through the stomach and small intestine every 90 minutes in a person who has fasted. Motilin secretion is inhibited after ingestion of food by mechanisms that are not fully understood.