Central Nervous System
BrainBrain
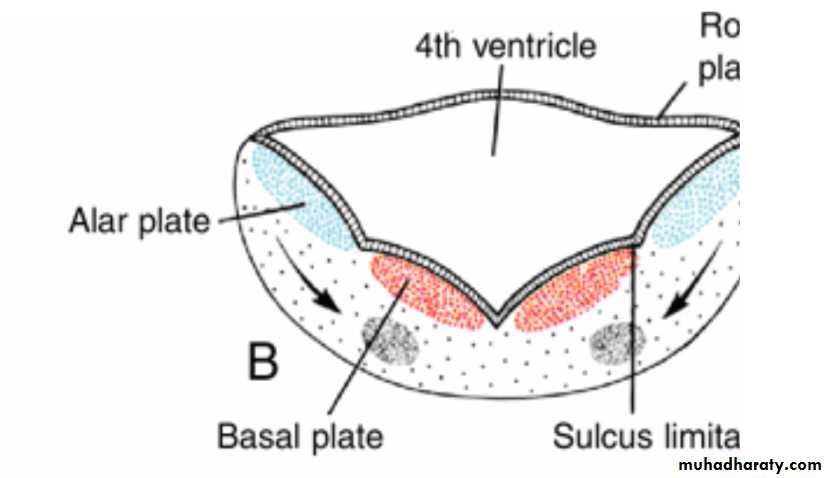
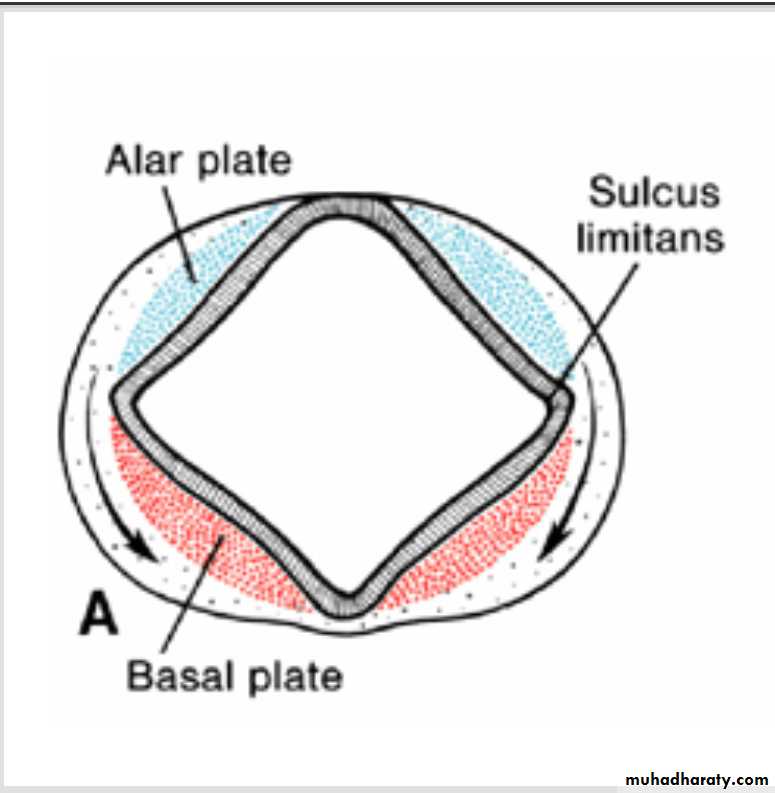
Distinct basal and Alar plates, representing motor and sensory areas, respectively, are found on each side of the midline in the rhombencephalon and mesencephalon.In the prosencephalon, however, the alar plates are accentuated and the basal plates regress.
Rhombencephalon: Hindbrain consists of the
(a) Myelencephalon, the most caudal of the brain vesicles, and(b) Metencephalon, which extends from the pontine flexure to the rhombencephalic isthmus.
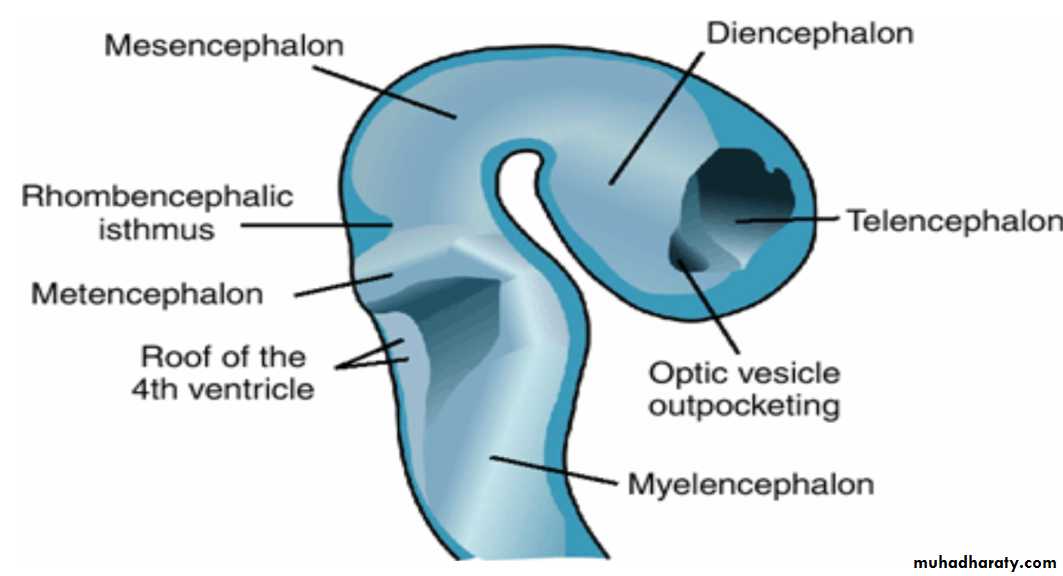
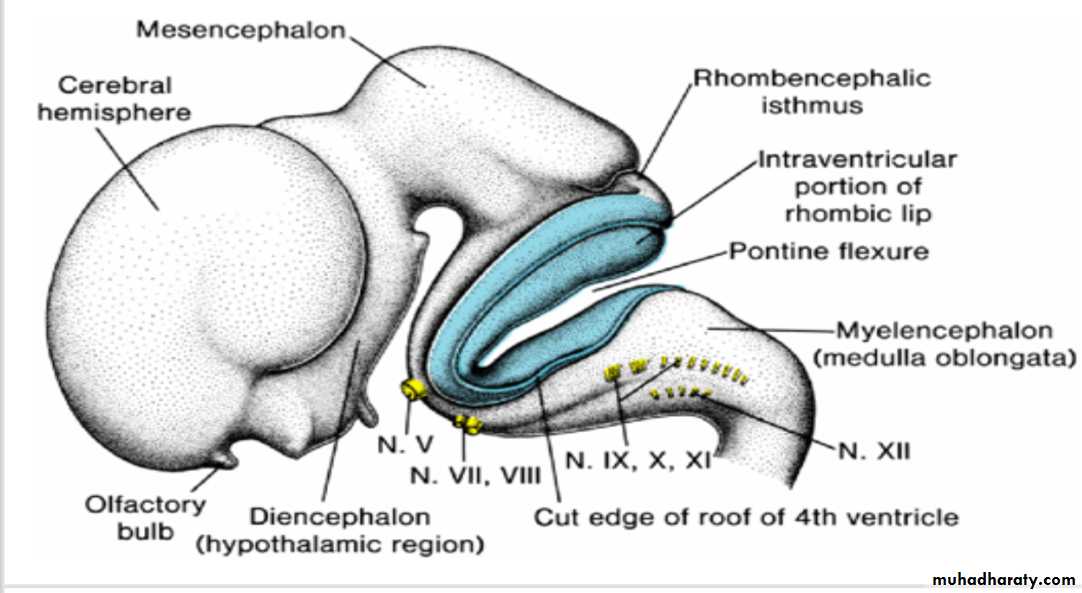
Rhombencephalon
MyelencephalonMyelencephalon
is a brain vesicle that gives rise to the medulla oblongata.
It differs from the spinal cord in that its lateral walls are everted .
Alar and basal plates separated by the sulcus limitans can be clearly distinguished.
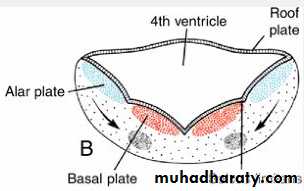
Myelencephalon
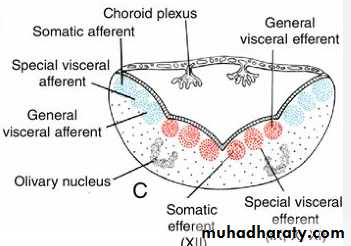
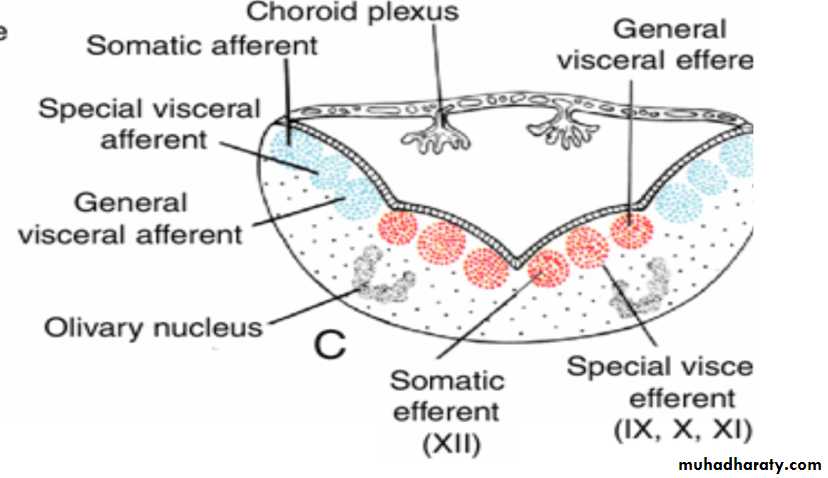
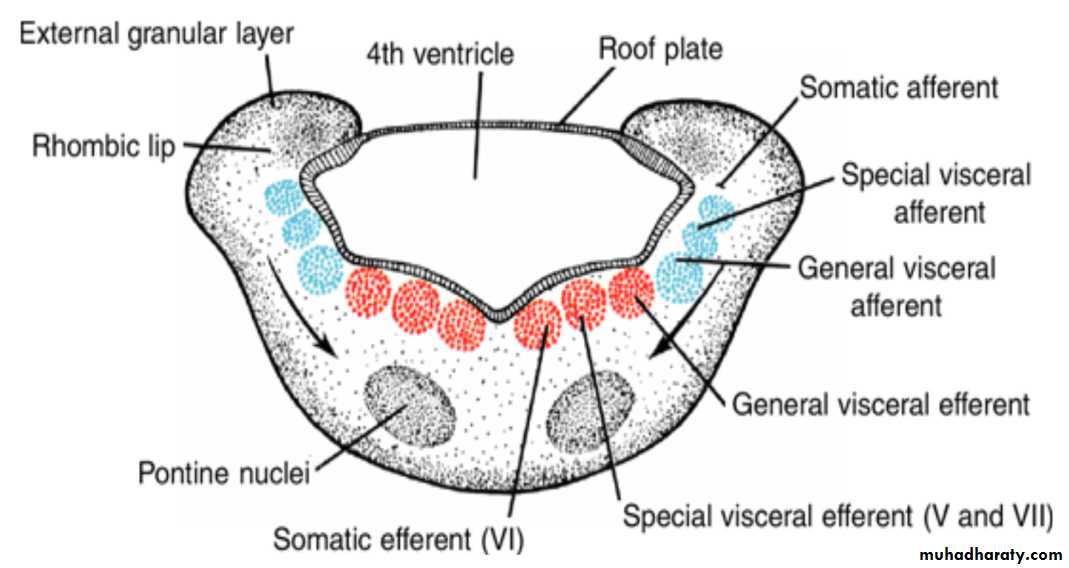
The basal plate, similar to that of the spinal cord, contains motor nuclei.These nuclei are divided into three groups: (a) a medial somatic efferent group,
(b) an intermediate special visceral efferent group, and
(c) a lateral general visceral efferent group.
(GIT HRT)
Somatic efferent groupThe first group contains motor neurons, which form the cephalic continuation of the anterior horn cells.
In the myelencephalon, it includes neurons of the hypoglossal nerve that supply the tongue musculature.
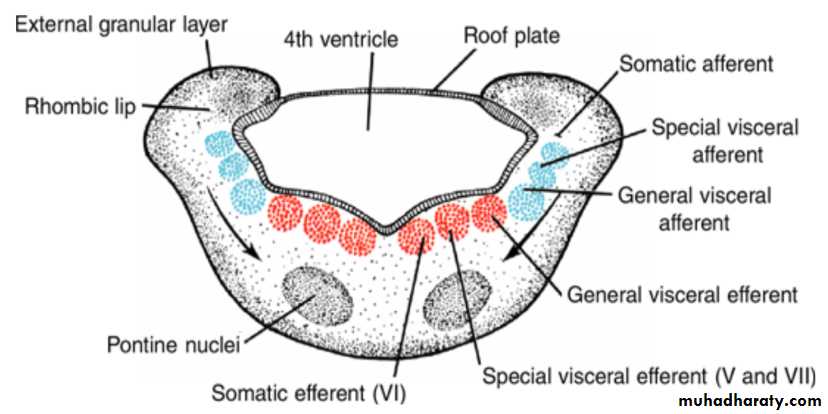
special visceral efferent group ( 12 9 10 11 10)
extends into the metencephalon, forming the special visceral efferent motor column.Its motor neurons supply striated muscles of the pharyngeal arches.
In the myelencephalon the column is represented by neurons of the accessory, vagus, and glossopharyngeal nerves.
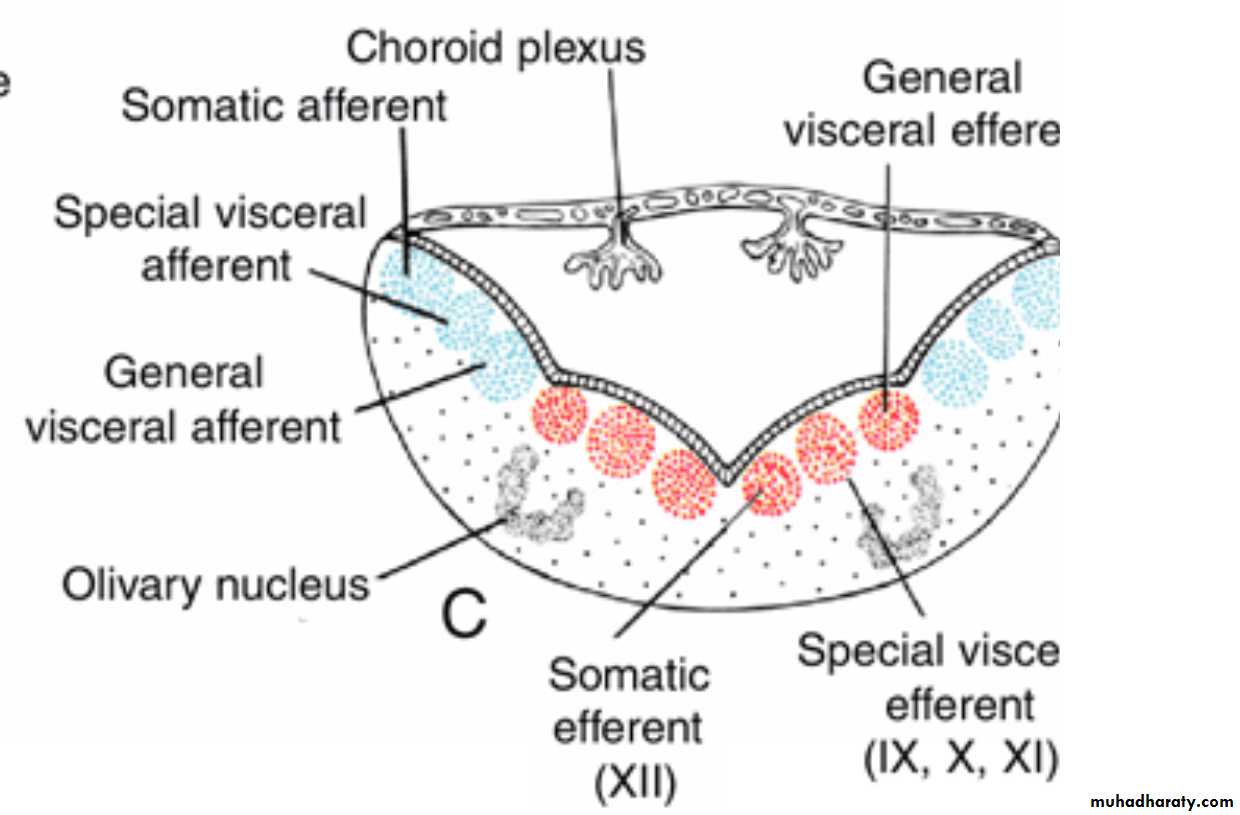
General visceral efferent group
contains motor neurons that supply involuntary musculature of the respiratory tract, intestinal tract, and heart.The alar plate contains three groups of sensory relay nuclei (851010)
The most lateral of these,the somatic afferent sensory group, receives impulses from the ear and surface of the head by way of the vestibulocochlear and trigeminal nerves.
The intermediate, or special visceral afferent, group receives impulses from taste buds of the tongue and from the palate, oropharynx, and epiglottis.
The medial, or general visceral afferent, group receives interoceptive information from the gastrointestinal tract and heart.
The roof plate of the myelencephalon consists of
a single layer of ependymal cells covered by vascular mesenchyme,the pia mater. The two combined are known as the tela choroidea.
Because of active proliferation of the vascular mesenchyme, a number of saclike invaginations project into the underlying ventricular cavity.
These tuft-like invaginations form the choroid plexus, which produces cerebrospinal fluid.
Arrows, path followed by cells of the alar plate to the olivary nuclear complex.
Rhombencephalon
MetencephalonMetencephalon
similar to the myelencephalon,
is characterized by basal and alar plates.Two new components form:
(a) the cerebellum; and (b) the pons,
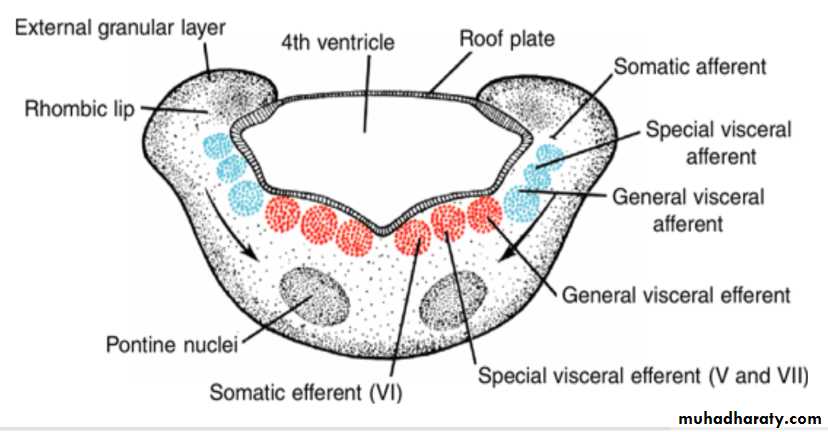
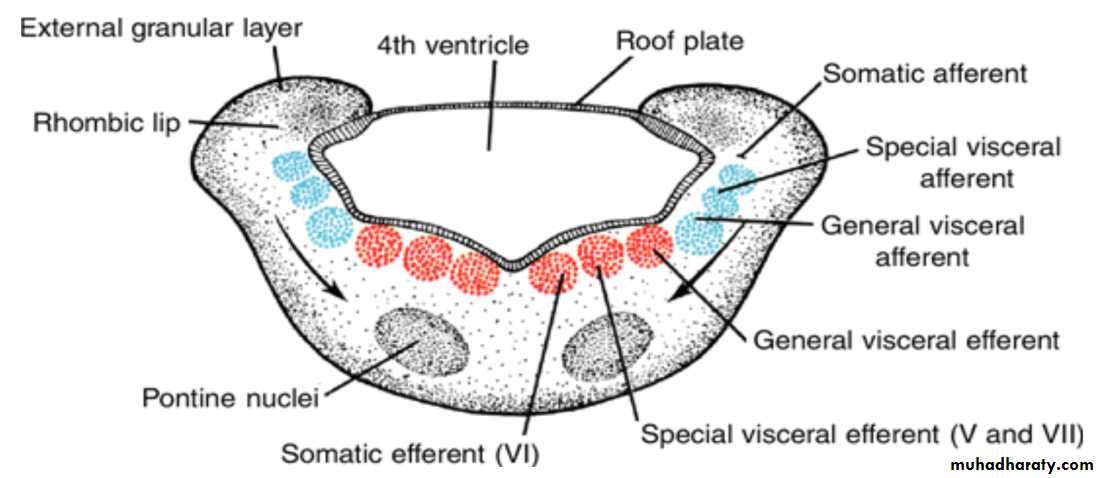
Each basal plate of the metencephaloncontains three groups of motor neurons:
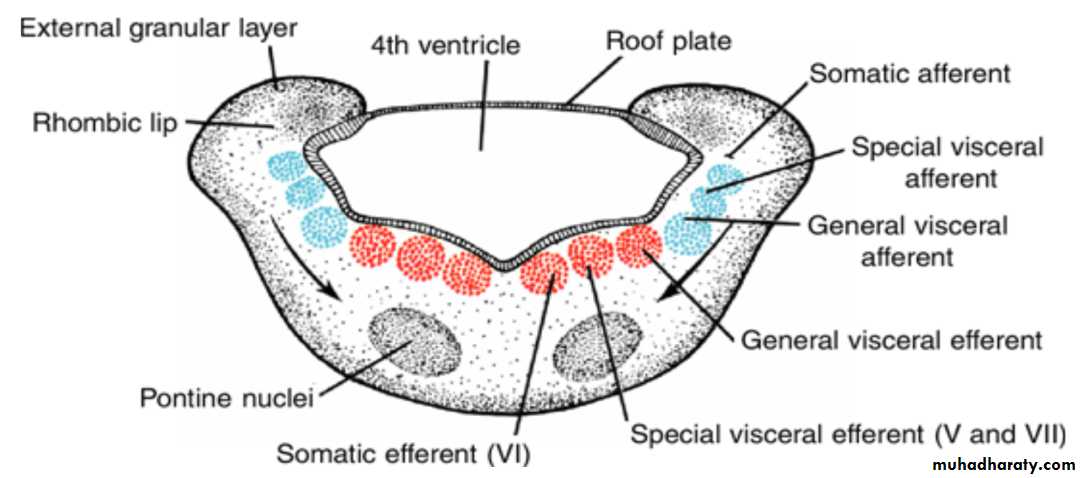
(a) the medial somatic efferent group, which gives rise to the nucleus of the abducens nerve
) b (the special visceral efferent group, containing nuclei of the trigeminal and facial nerves, which innervate the musculature of the first and second pharyngeal arches; and
(c) the general visceral efferent group, whose axons supply the submandibular and sublingual glands.
The alar plates of the metencephalon
contain three groups of sensory nuclei:
(a) a lateral somatic afferent group, which contains neurons of the trigeminal nerve and a small portion of the vestibulocochlear complex(b) the special visceral afferent group, and
(c the general visceral afferent group
Metencephalon
The marginal layer of the basal plates of the metencephalon expands as it makes a bridge for nerve fibers connecting the cerebral cortex and cerebellar cortex with the spinal cord.Hence this portion of the metencephalon is known as the pons (bridge).
In addition to nerve fibers, the pons contains the pontine nuclei, which originate in the alar plates of the metencephalon and myelencephalon, arrows.
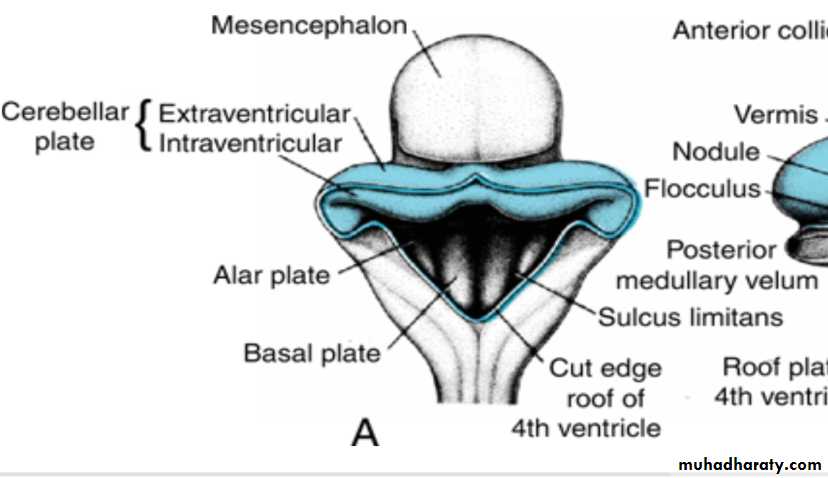
Cerebellum
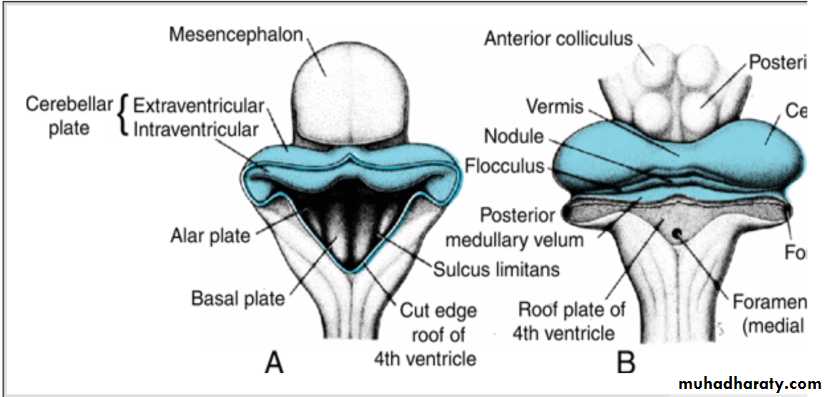
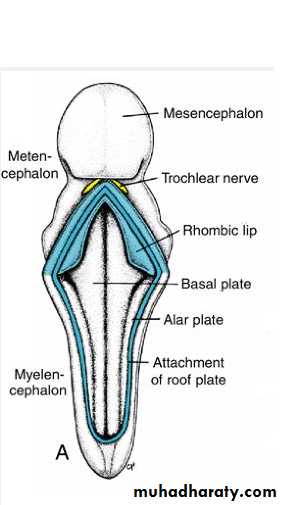
The dorsolateral parts of the alar plates bend medially and form the rhombic lips.In the caudal portion of the metencephalon, the rhombic lips are widely separated, but immediately below the mesencephalon they approach each other in the midline .
As a result of a further deepening of the pontine flexure, the rhombic lips compress cephalocaudally and form the cerebellar plate.
The cerebellar plate
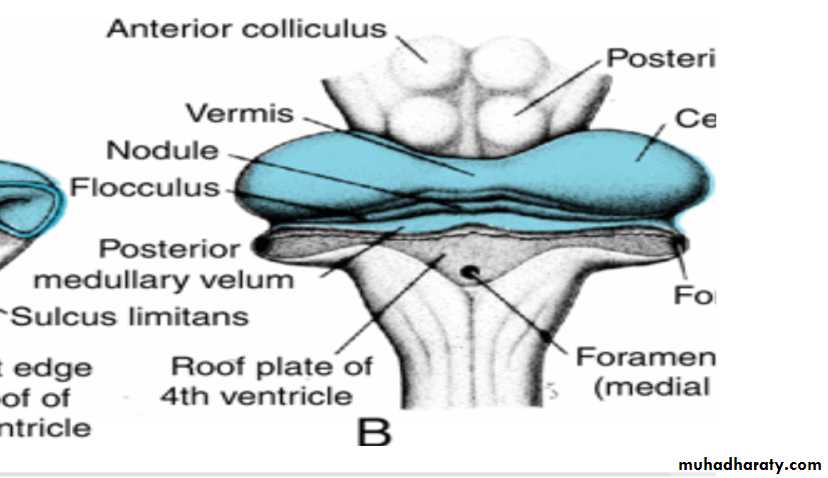
In a 12-week embryo, this plate shows a small midline portion, the vermis, and two lateral portions, the hemispheres.
A transverse fissure soon separates the nodule from the vermis and the lateral flocculus from the hemispheres .
This flocculonodular lobe is phylogenetically the most primitive part of the cerebellum.
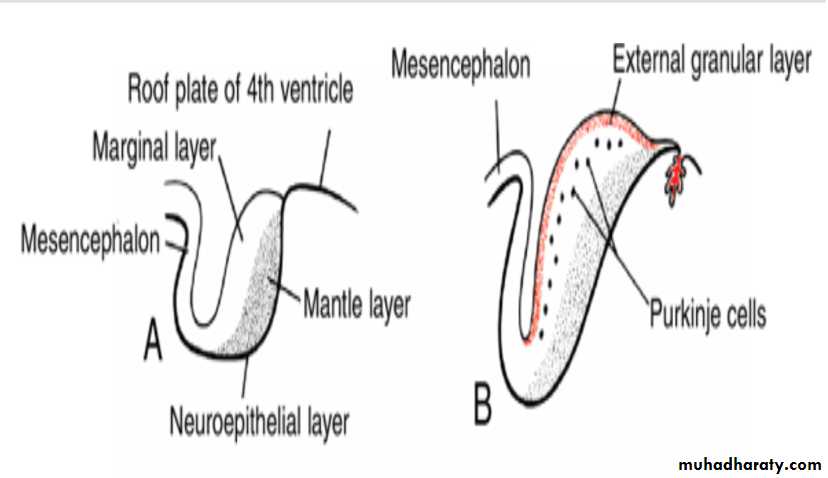
The cerebellar plate
Initially, the cerebellar plate consists of neuroepithelial, mantle, and marginal layers (A).During further development, a number of cells formed by the neuroepithelium migrate to the surface of the cerebellum to form the external granular layer. Cells of this layer retain their ability to divide and form a proliferative zone on the surface of the cerebellum (B,C).
B(12w), C(13w),D(15w)
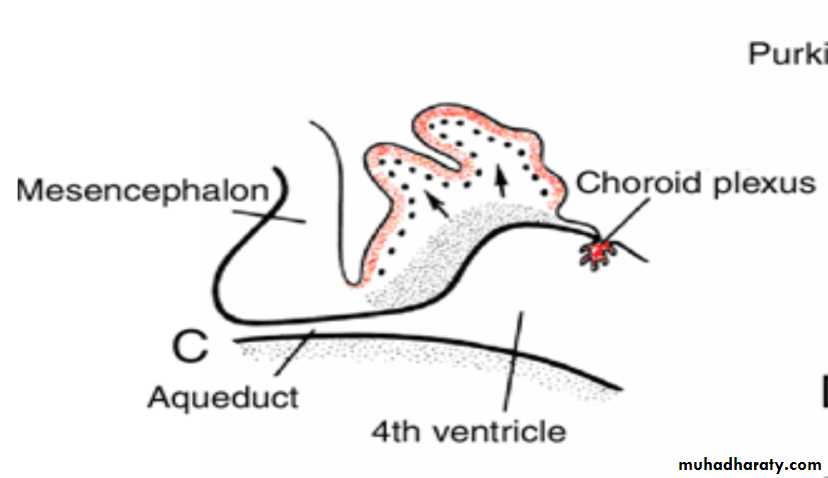
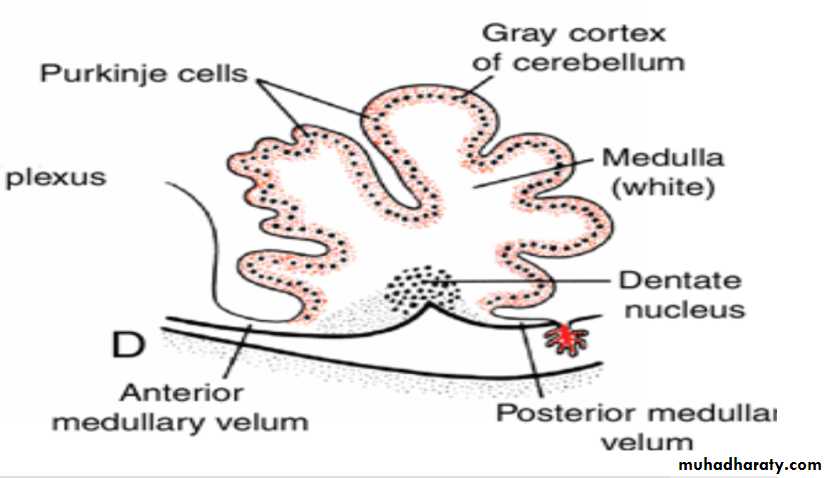
Cerebellum
In the sixth month of development, the external granular layer gives rise to various cell types. (granule, Basket and stellate cells)The cortex of the cerebellum, consisting of Purkinje cells, Golgi II neurons, and neurons produced by the external granular layer, reaches its definitive size after birth
The deep cerebellar nuclei, such as the dentate nucleus, reach their final position before birth.
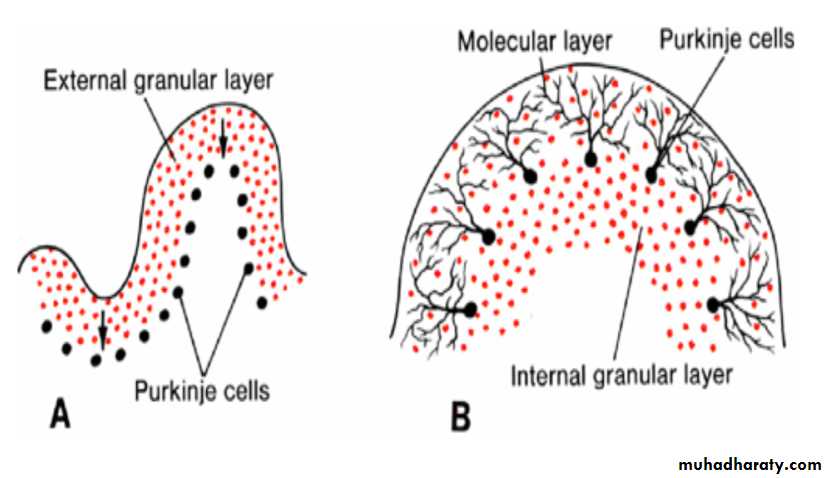
Mesencephalon
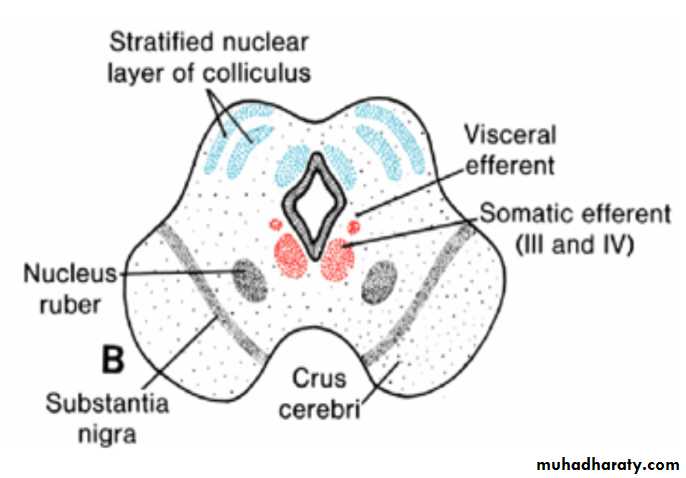
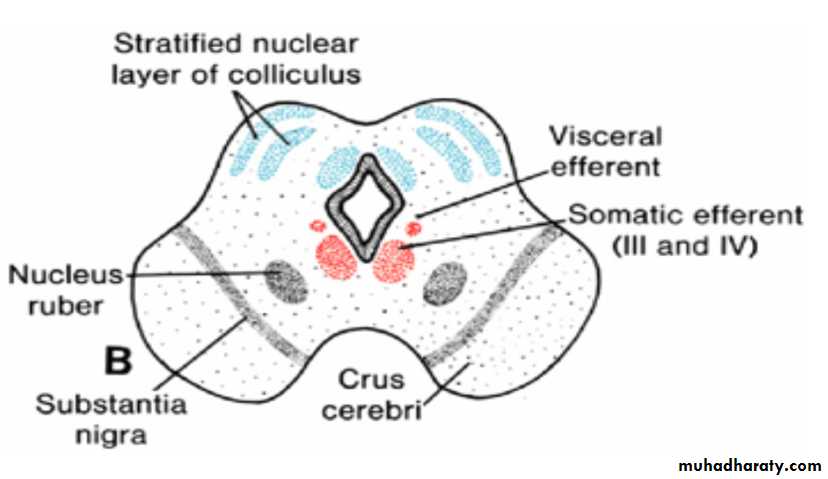
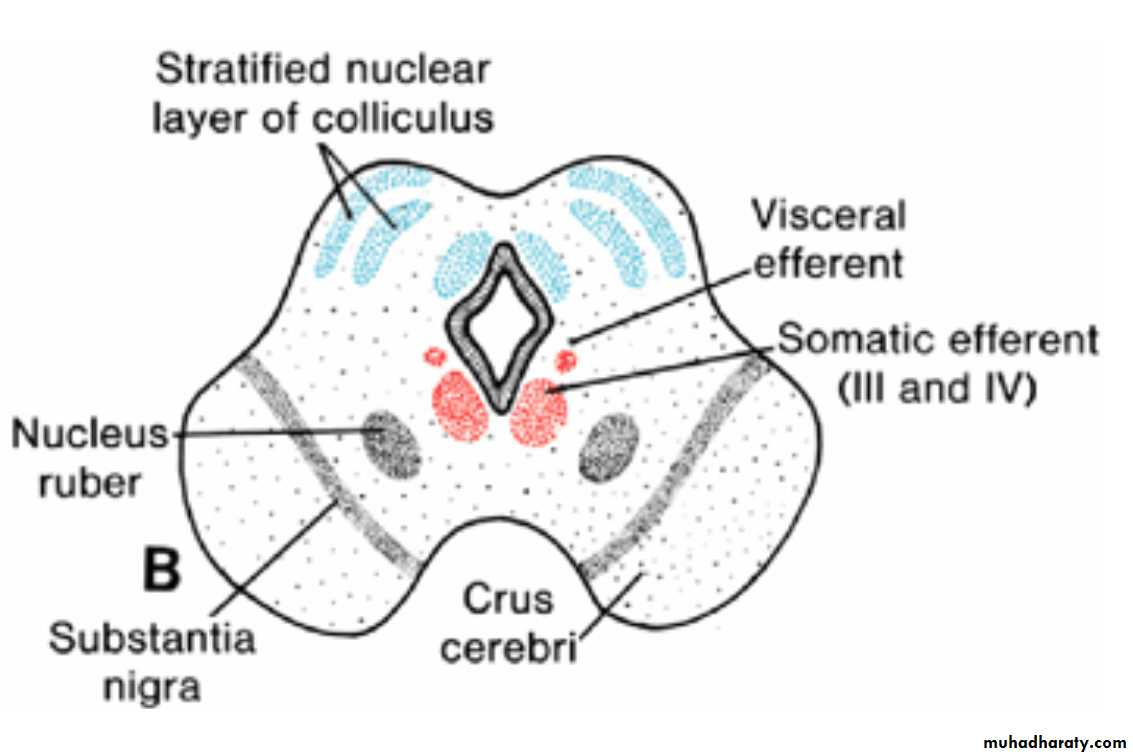
Mesencephalon: Midbraineach basal plate contains two groups of motor nuclei:
(a) a medial somatic efferent group, represented by the oculomotor and trochlear nerves, which innervate the eye musculature; and
(b) a small general visceral efferent group, represented by the nucleus of Edinger-Westphal, which innervates the sphincter pupillary muscle
The marginal layer of each basal plate enlarges and forms the crus cerebri. These crura serve as pathways for nerve fibers descending from the cerebral cortex to lower centers in the pons and spinal cord.
Arrows in A indicate the path followed by cells of the alar plate to form the nucleus ruber and substantia nigra.
Mesencephalon: Midbrain the alar plates
Initially, the alar plates of the mesencephalon appear as two longitudinal elevations separated by a shallow midline depression
With further development, a transverse groove divides each elevation into an anterior (superior) and a posterior (inferior) colliculus.
The posterior colliculi serve as synaptic relay stations for auditory reflexes; the anterior colliculi function as correlation and reflex centers for visual impulses.
The colliculi are formed by waves of neuroblasts migrating into the overlying marginal zone. Here they are arranged in layers.
Prosencephalon(Forebrain)
Prosencephalon: Forebrain consists ofthe Telencephalon, which forms the cerebral hemispheres, and
the Diencephalon, which forms the optic cup and stalk, pituitary, thalamus, hypothalamus, and epiphysis.
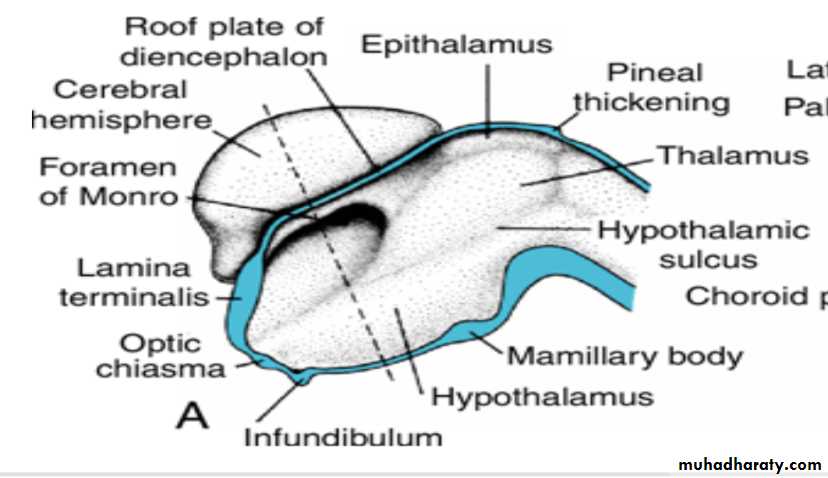
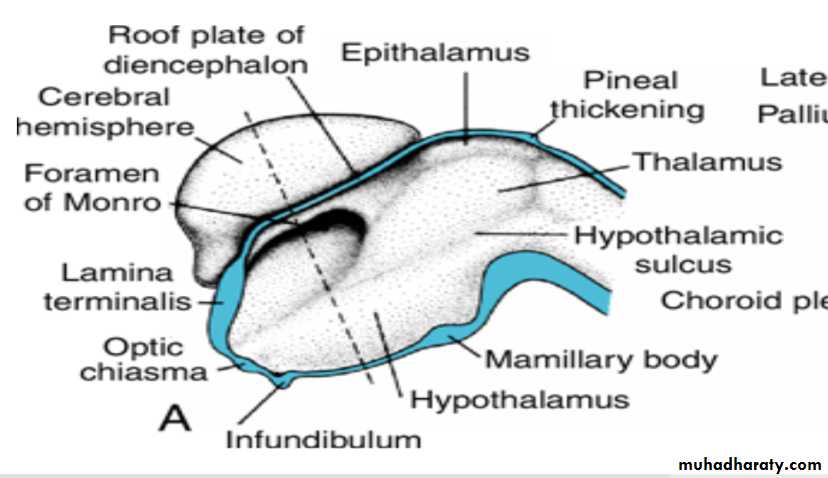
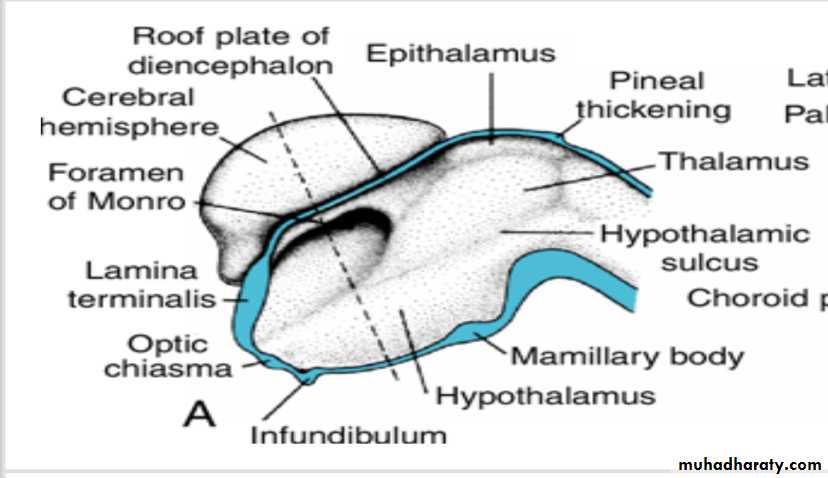
Diencephalon
develops from the median portion of the prosencephalonis thought to consist of a roof plate and two alar plates but to lack floor and basal plates.
The roof plate of the diencephalon choroid plexus of the third ventricle
consists of a single layer of ependymal cells covered by vascular mesenchyme.
Together, these layers give rise to the choroid plexus of the third ventricle.The roof plate of the diencephalon pineal body
The most caudal part of the roof plate develops into the pineal body, or epiphysis.
This body initially appears as an epithelial thickening in the midline, but by the seventh week it begins to evaginateEventually, it becomes a solid organ on the roof of the mesencephalon
The roof plate of the diencephalon pineal body
serves as a channel through which light and darkness affect endocrine and behavioral rhythms.In the adult, calcium is frequently deposited in the epiphysis and then serves as a landmark on radiographs of the skull.
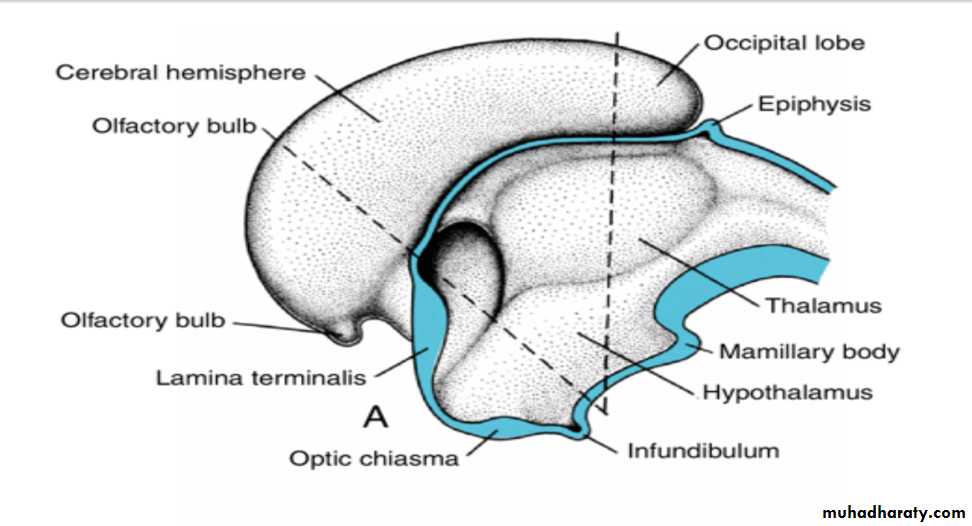
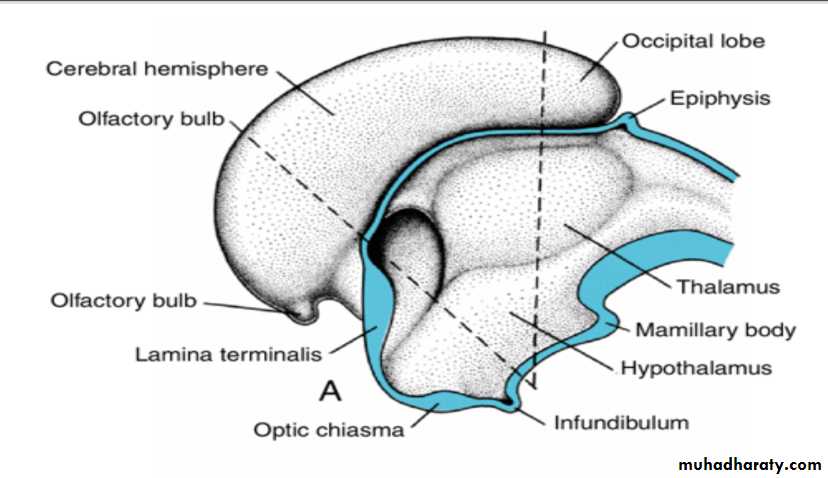
Alar Plate, Thalamus, and Hypothalamus
The alar plates form the lateral walls of the diencephalon.A groove, the hypothalamic sulcus, divides the plate into a dorsal and a ventral region, the thalamus and hypothalamus, respectively .
As a result of proliferative activity, the thalamus gradually projects into the lumen of the diencephalon. Frequently this expansion is so great that thalamic regions from the right and left sides fuse in the midline, forming the massa intermedia, or interthalamic connexus.
Alar plate Hypothalamus
The hypothalamus, forming the lower portion of the alar plate, differentiates into a number of nuclear areas that regulate the visceral functions, including sleep, digestion, body temperature, and emotional behavior.
One of these groups, the mamillary body, forms a distinct protuberance on the ventral surface of the hypothalamus on each side of the midline.
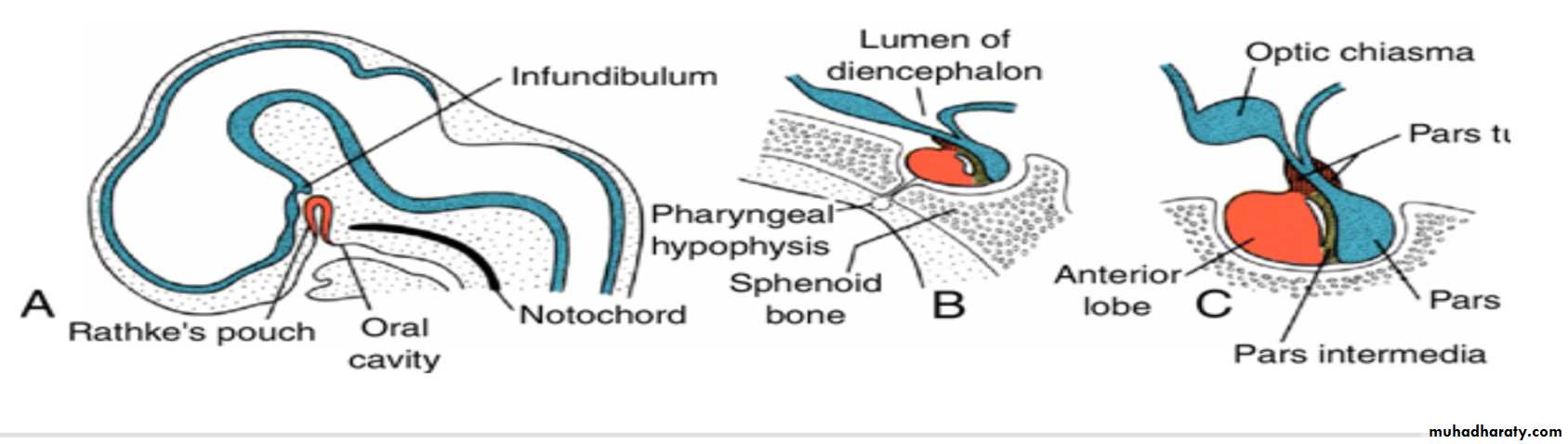
Hypophysis or Pituitary Gland develops from two completely different parts
(a (Rathke's pouch :an ectodermal outpocketing of the stomodeum immediately in front of the buccopharyngeal membrane(b) the infundibulum :a downward extension of the diencephalon,.
Hypophysis or Pituitary Gland
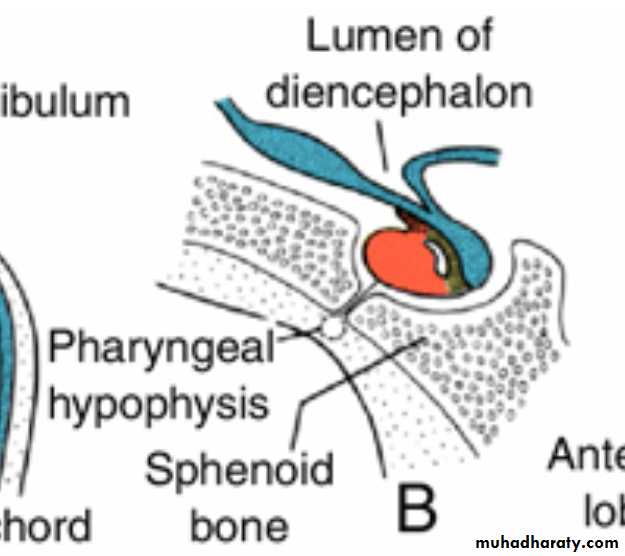
When the embryo is approximately 3 weeks old, Rathke's pouch appears as an evagination of the oral cavity and subsequently grows dorsally toward the infundibulum.By the end of the second month, it loses its connection with the oral cavity and is then in close contact with the infundibulum.
Hypophysis or Pituitary Gland
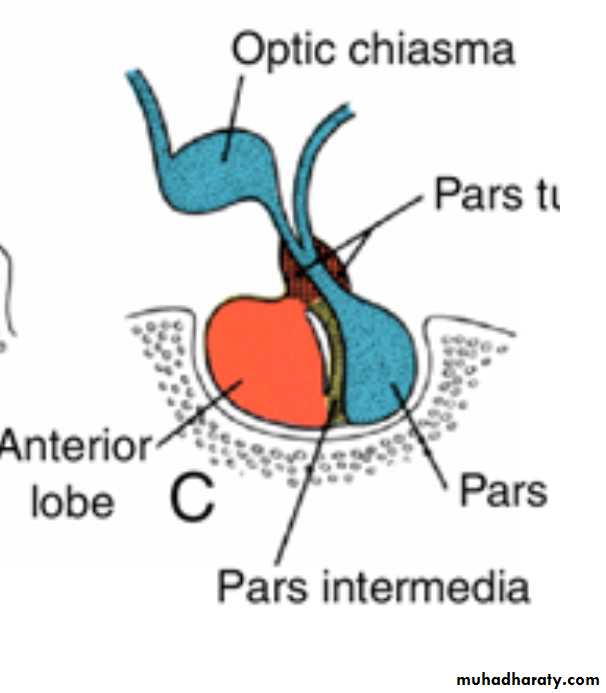
cells in the anterior wall of Rathke's pouch increase rapidly in number and form the anterior lobe of the hypophysis, or Adenohypophysis .
A small extension of this lobe, the pars tuberalis, grows along the stalk of the infundibulum and eventually surrounds it
The posterior wall of Rathke's pouch develops into the pars intermedia, which in humans seems to have little significance.
The infundibulum gives rise to the stalk and the pars nervosa, or posterior lobe of the hypophysis( neurohypophysis). It is composed of neuroglial cells. In addition, it contains a number of nerve fibers from the hypothalamic area.
Clinical Correlates Hypophyseal Defects
Occasionally a small portion of Rathke's pouch persists in the roof of the pharynx as a pharyngeal hypophysis.Craniopharyngiomas arise from remnants of Rathke's pouch. They may form within the sella turcica or along the stalk of the pituitary but usually lie above the sella.
They may cause hydrocephalus and pituitary dysfunction (e.g., diabetes insipidus, growth failure).
Telencephalon
Telencephalonthe most rostral of the brain vesicles,
consists of two lateral outpocketings, the cerebral hemispheres, and a median portion, the lamina terminalis.
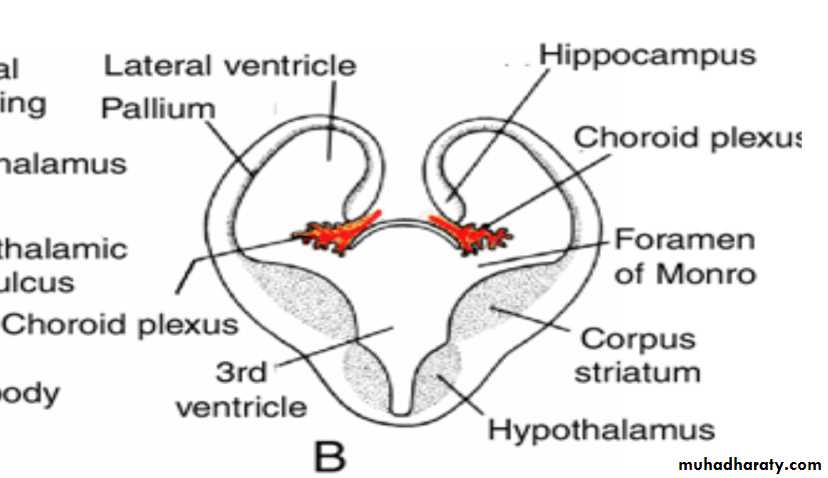
The cavities of the hemispheres, the lateral ventricles, communicate with the lumen of the diencephalon through the interventricular foramina of Monro.
Cerebral Hemispheres
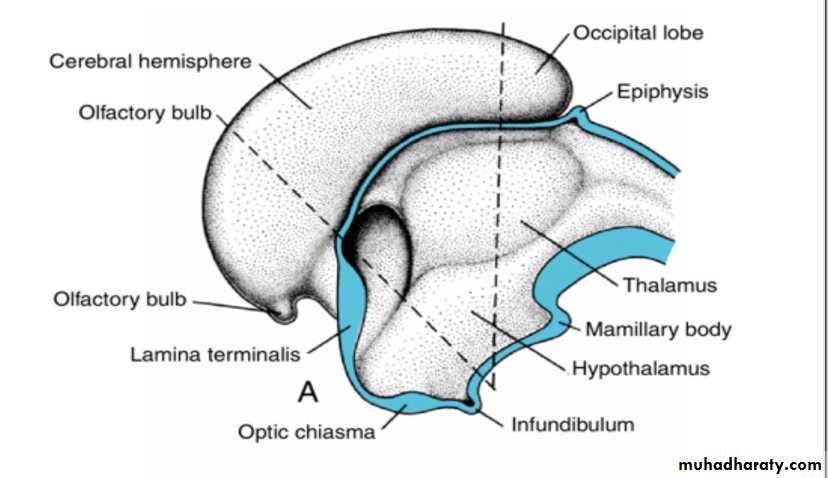
The cerebral hemispheres arise at the beginning of the fifth week of development as bilateral evaginations of the lateral wall of the prosencephalon.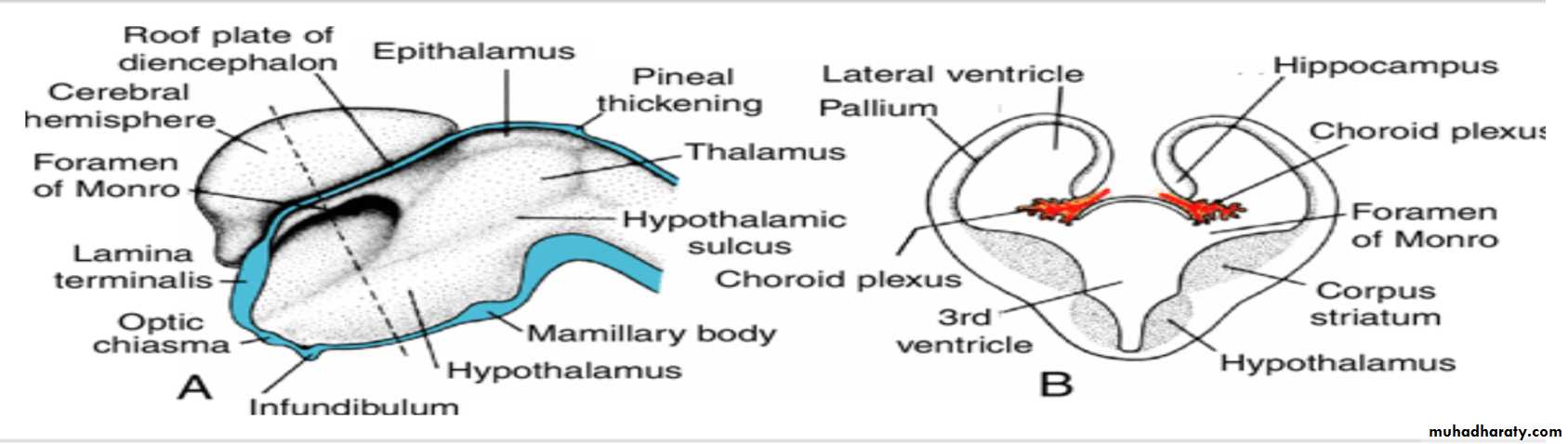
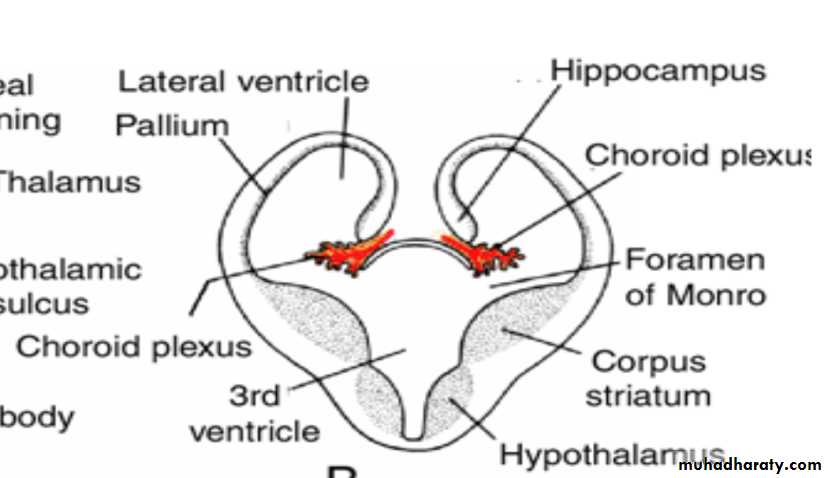
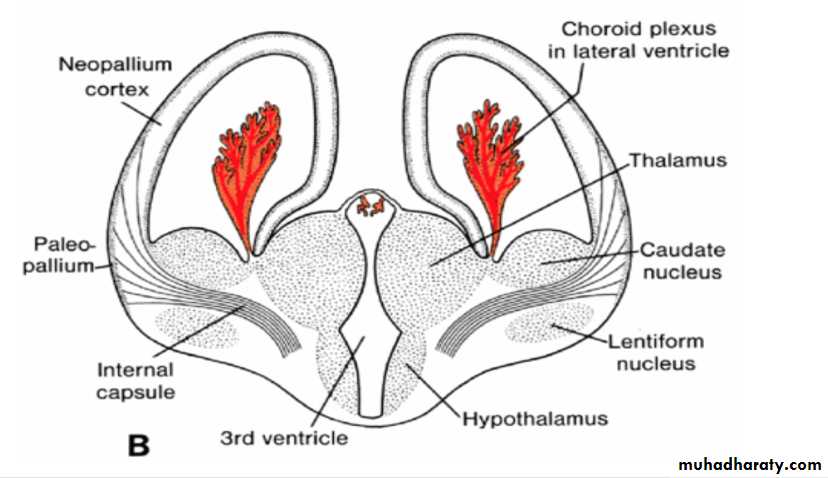
Corpus striatum
By the middle of the second month the basal part of the hemispheres (i.e., the part that initially formed the forward extension of the thalamus( begins to grow and bulges into the lumen of the lateral ventricle and into the floor of the foramen of Monro (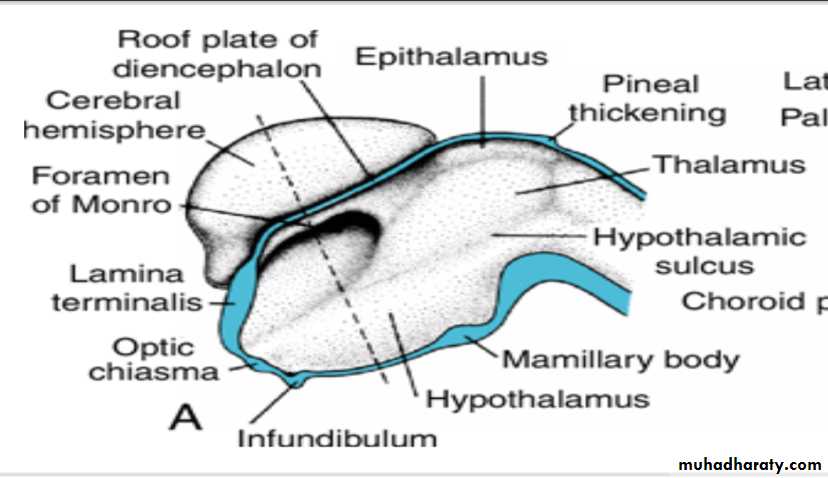
Corpus striatum
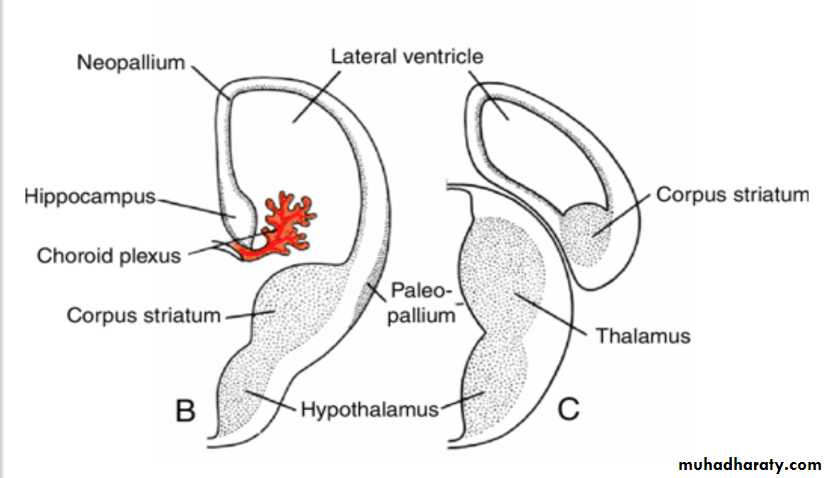
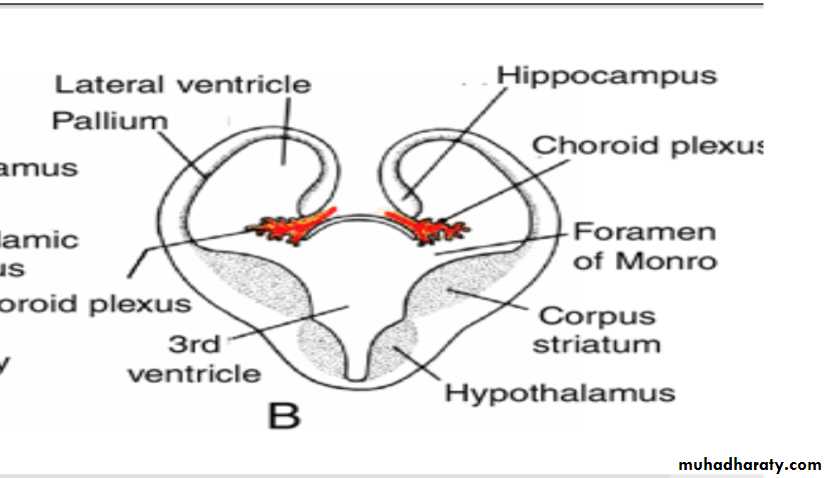
In transverse sections, the rapidly growing region has a striated appearance and is therefore known as the corpus striatum
B,C. Transverse sections through the right half of the telencephalon and diencephalon at the level of the broken lines in A.
Choroid plexus
In the region where the wall of the hemisphere is attached to the roof of the diencephalon, the wall fails to develop neuroblasts and remains very thin.
Here the hemisphere wall consists of a single layer of ependymal cells covered by vascular mesenchyme, and together they form the choroid plexus.
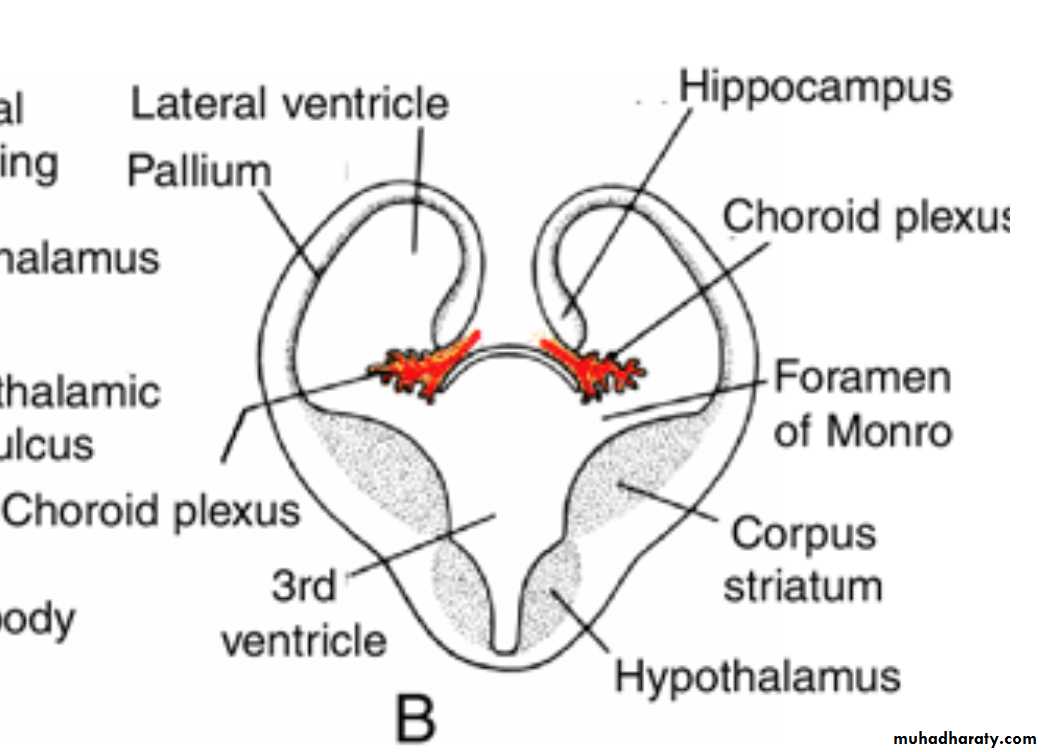
The choroid plexus
should have formed the roof of the hemisphere, but as a result of the disproportionate growth of the various parts of the hemisphere, it protrudes into the lateral ventricle along the choroidal fissure.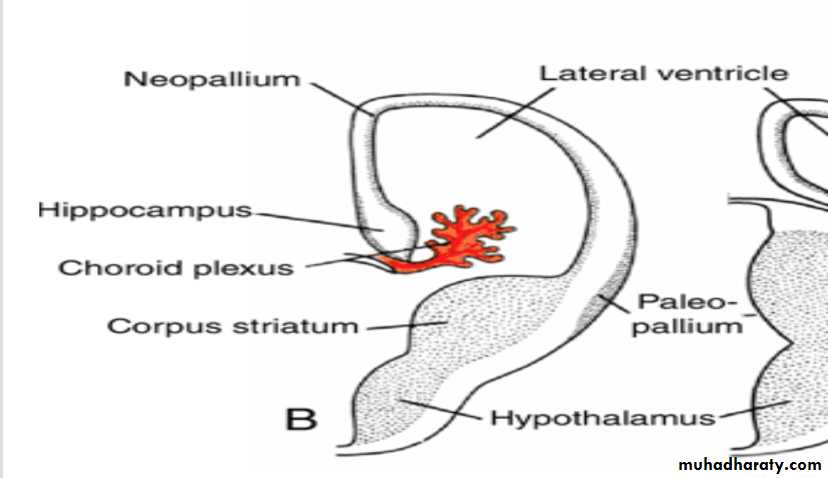
Hippocampus
Immediately above the choroidal fissure, the wall of the hemisphere thickens, forming the hippocampus .This structure, whose primary function is olfaction, bulges into the lateral ventricle.
With further expansion, the hemispheres cover the lateral aspect of the diencephalon, mesencephalon, and cephalic portion of the metencephalon.
Corpus striatum
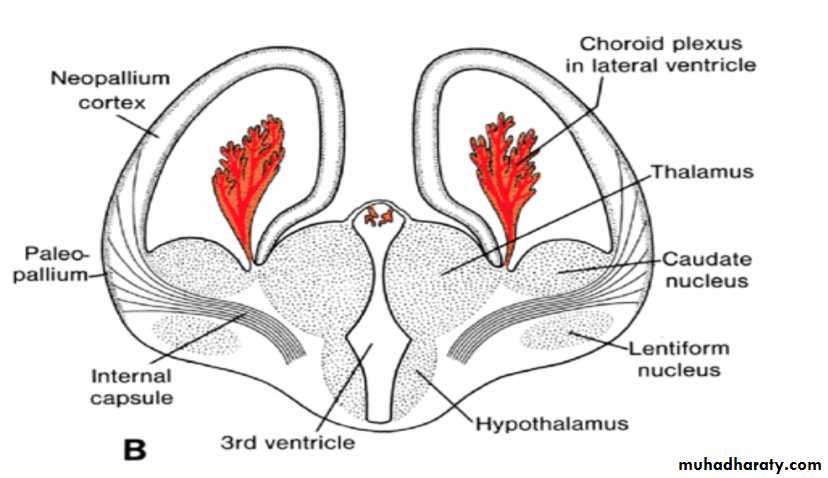
The corpus striatum, being a part of the wall of the hemisphere, expands posteriorly and is divided into two parts:(a) a dorsomedial portion, the caudate nucleus, and (b) a ventrolateral portion, the lentiform nucleus.
This division is accomplished by axons passing to and from the cortex of the hemisphere and breaking through the nuclear mass of the corpus striatum.
The fiber bundle thus formed is known as the internal capsule .
At the same time, the medial wall of the hemisphere and the lateral wall of the diencephalon fuse, and the caudate nucleus and thalamus come into close contact.
The cerebral hemispheres
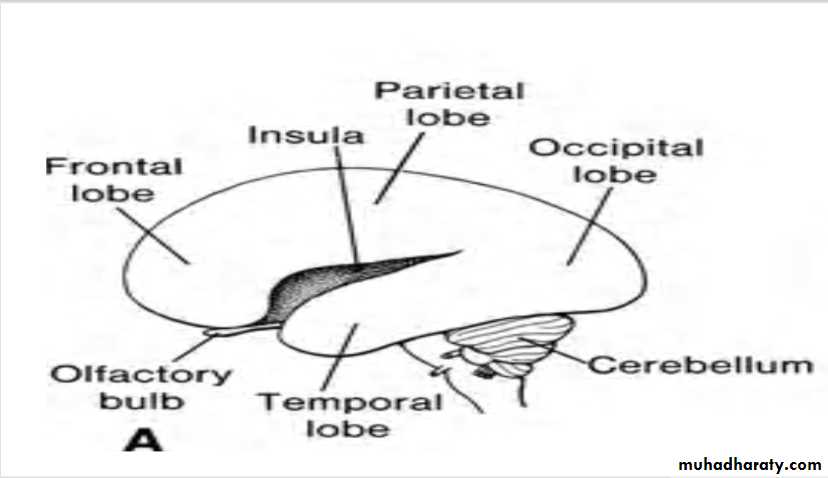
Continuous growth of the cerebral hemispheres in anterior, dorsal, and inferior directions results in the formation of frontal, temporal, and occipital lobes, respectively.As growth in the region overlying the corpus striatum slows, however, the area between the frontal and temporal lobes becomes depressed and is known as the insula.
This region is later overgrown by the adjacent lobes and at the time of birth is almost completely covered.
During the final part of fetal life, the surface of the cerebral hemispheres grows so rapidly that a great many convolutions (gyri( separated by fissures and sulci appear on its surface.
Cortex Development
At birth, the cortex has a stratified appearance due to differentiation of the cells in layers.The motor cortex contains a large number of pyramidal cells, and the sensory areas are characterized bygranular cells.
Olfactory Bulbs
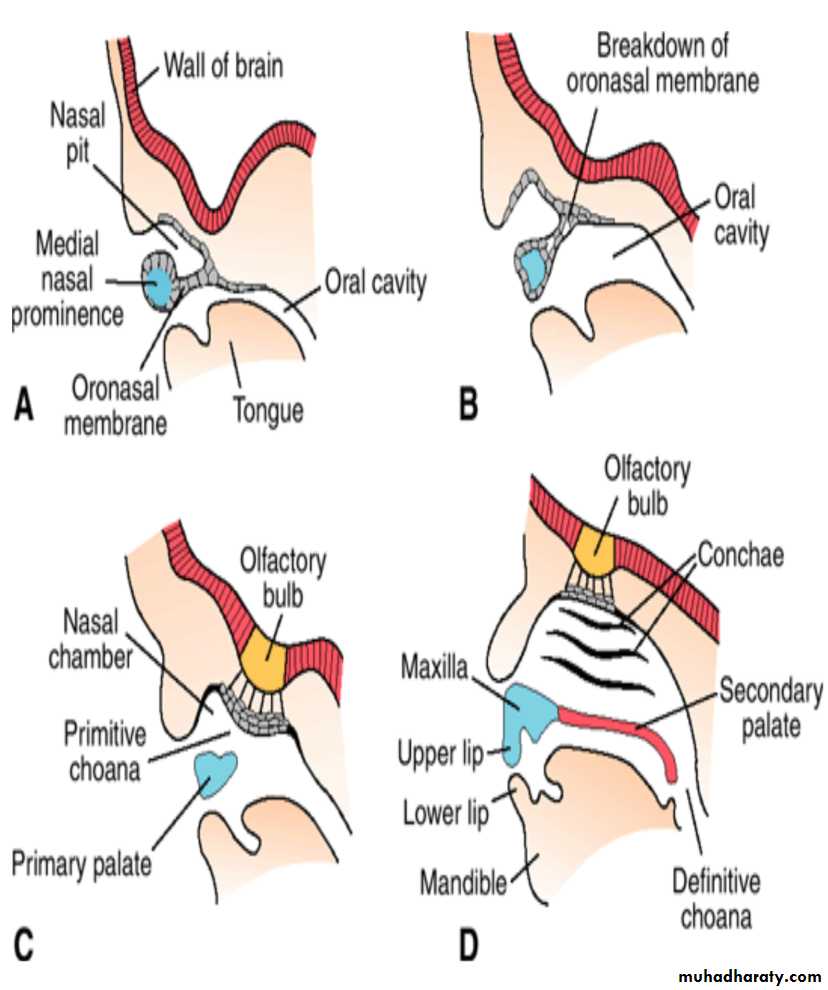
Differentiation of the olfactory system is dependent on epithelial– mesenchymal interactions.These occur between neural crest cells and ectoderm of the frontonasal prominence to form the olfactory placodes and between these same crest cells and the floor of the telencephalon to form the olfactory bulbs.
The olfactory nerve
Cells in the nasal placodes differentiate into primary sensory neurons of the nasal epithelium whose axons grow and make contact with secondary neurons in the developing olfactory bulbs.By the seventh week, these contacts are well established.
As growth of the brain continues, the olfactory bulbs and the olfactory tracts of the secondary neurons lengthen, and together they constitute the olfactory nerve.
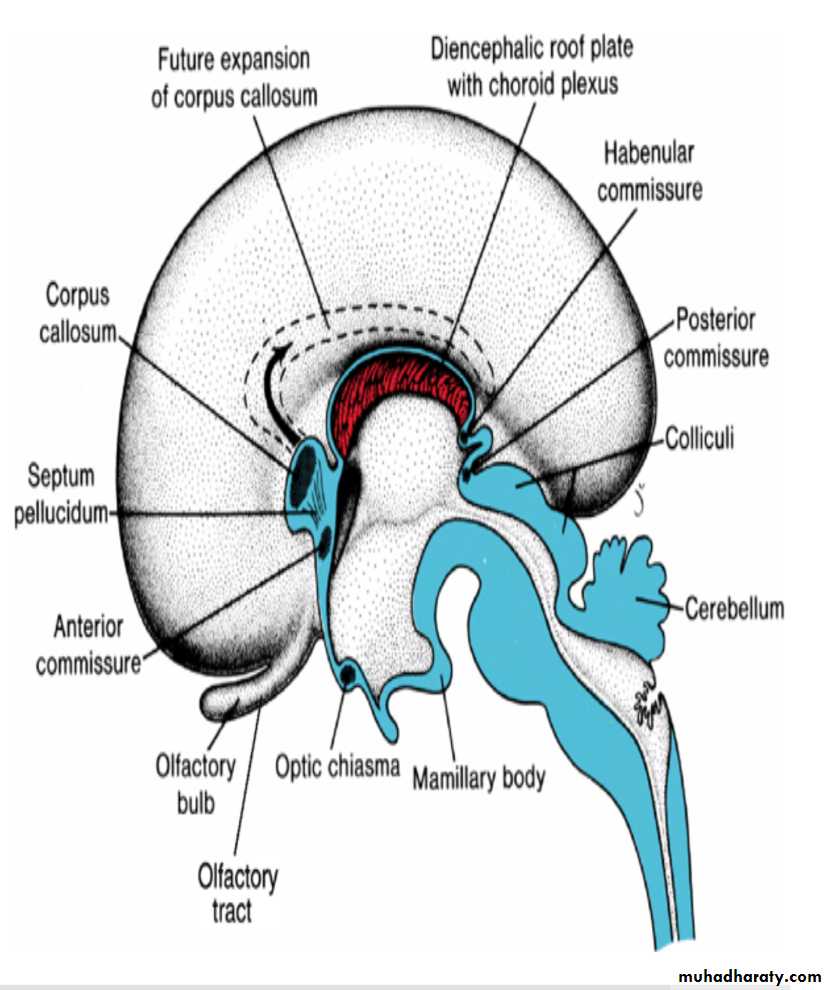
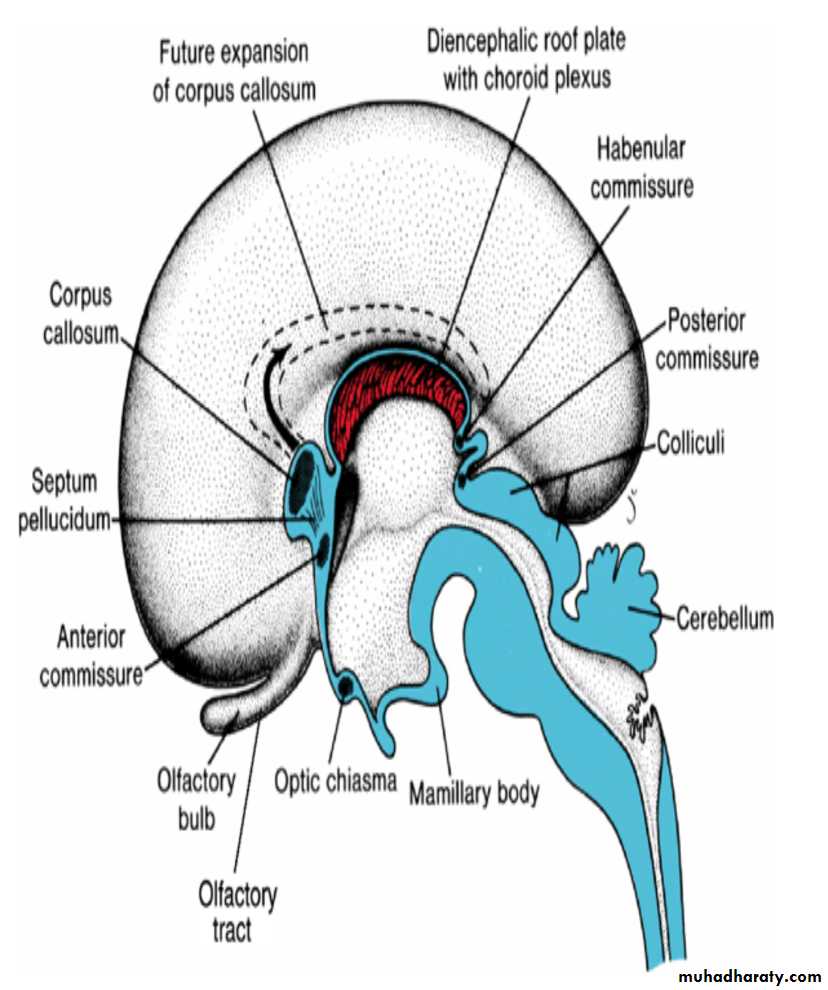
Commissures
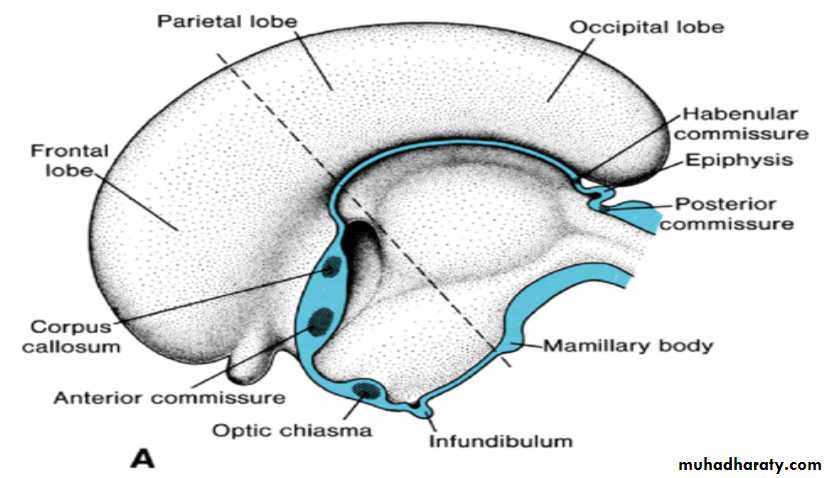
In the adult, a number of fiber bundles, the commissures, which cross the midline, connect the right and left halves of the hemispheres.The most important fiber bundles make use of the lamina terminalis
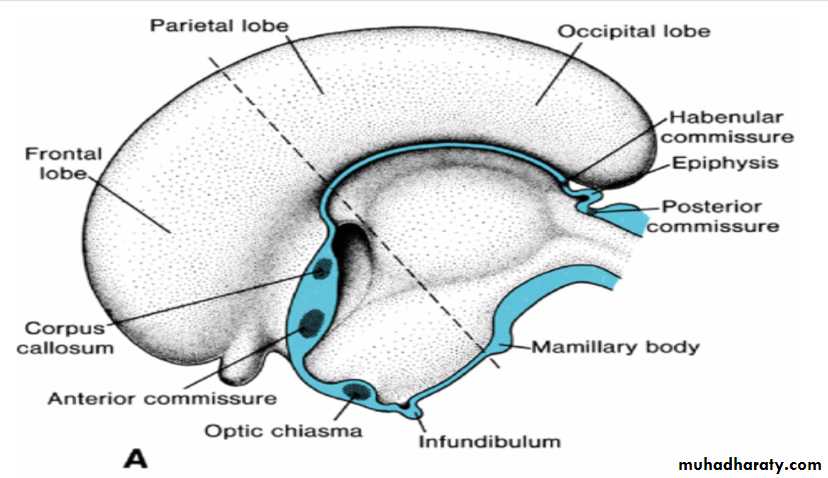
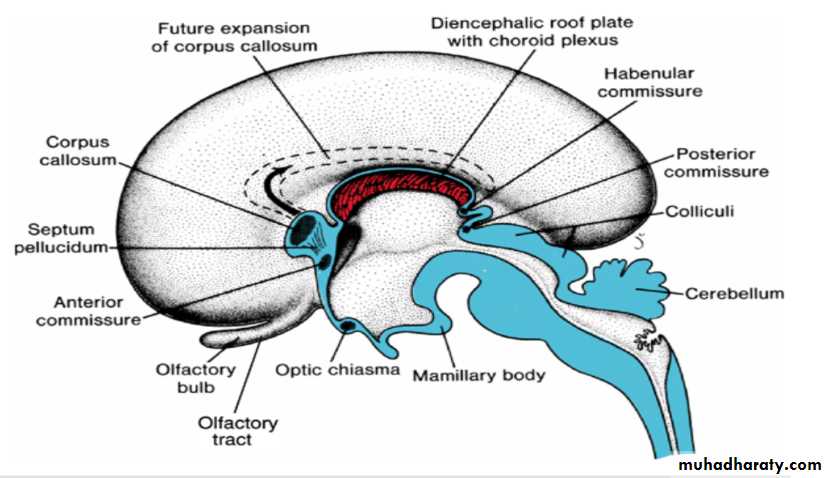
Commissures the anterior commissure & the hippocampal commissure
The first of the crossing bundles to appear is the anterior commissure. It consists of fibers connecting the olfactory bulb and related brain areas of one hemisphere to those of the opposite side.
The second commissure to appear is the hippocampal commissure, or fornix commissure. Its fibers arise in the hippocampus and converge on the lamina terminalis close to the roof plate of the diencephalon. From here the fibers continue, forming an arching system immediately outside the choroid fissure, to the mamillary body and the hypothalamus.
Commissure the corpus callosum
The most important commissure is the corpus callosum. It appears by the 10th week of development and connects the non olfactory areas of the right and the left cerebral cortex.Initially, it forms a small bundle in the lamina terminalis. As a result of continuous expansion of the cerbral cortex , it extends first anteriorly and then posteriorly, arching over the thin roof of the diencephalon
Commissures
In addition to the three commissures developing in the lamina terminalis, three more appear.
Two of these, the posterior and habenular commissures, are just below and rostral to the stalk of the pineal gland.
The third, the optic chiasma, which appears in the rostral wall of the diencephalon, contains fibers from the medial halves of the retinae.
Cranial Nerves
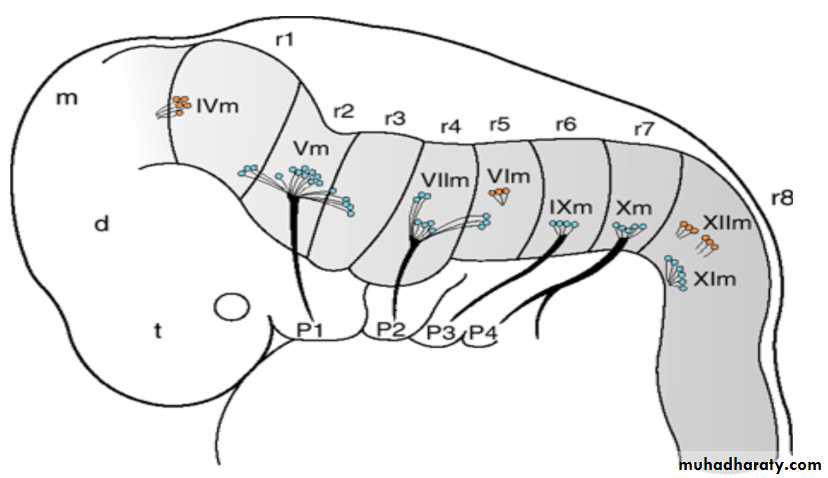
By the fourth week of development, nuclei for all 12 cranial nerves are present.All except the olfactory (I) and optic (II) nerves arise from the brainstem, and of these only the oculomotor (III) arises outside the region of the hindbrain.
The hindbrain is divided into eight rhombomeres (r1–r8), and these structures give rise to to motor nuclei of cranial nerves IV, V, VI, VII, IX, X, XI, and XII.
Establishment of this segmental pattern appears to be directed by mesoderm collected into somitomeres beneath the overlying neuroepithelium.
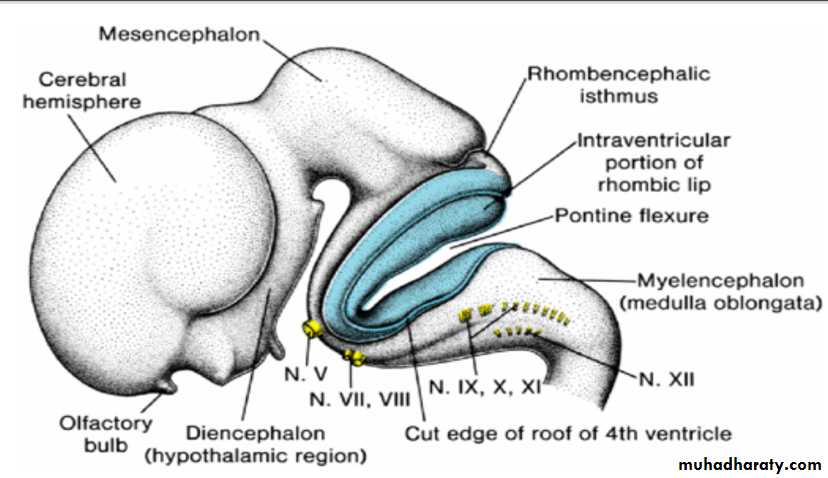
Cranial nerve sensory ganglia
Motor neurons for cranial nuclei are within the brainstem, while sensory ganglia are outside of the brain. Thus, the organization of cranial nerves is homologous to that of spinal nerves, although not all cranial nerves contain both motor and sensory fibers .Cranial nerve sensory ganglia originate from ectodermal placodes and neural crest cells. Ectodermal placodes include the nasal, otic, and four Epibranchial placodes represented by ectodermal thickenings dorsal to the pharyngeal (branchial) arches .
Epibranchial placodes contribute to ganglia for nerves of the pharyngeal arches (V, VII, IX, and X). Parasympathetic (visceral efferent) ganglia are derived from neural crest cells, and their fibers are carried by cranial nerves III, VII, IX, and X